|
●何故、生体膜研究が重要か?
ヒトのゲノムは30億塩基対から成る。2003年、ヒトのゲノムの全塩基配列が決定された。
これは、ちょうど生命を解く暗号を手に入れたことになる。
現在、この塩基配列上のどの部分がタンパク質やRNAをコードしている領域か、
どの部分が遺伝子の発現を制御している領域かといった注釈をつける作業
(annotattion)が進められている。
しかしながら、たとえ全遺伝子に注釈がつけられても解決できない問題がある。
それは、我々の身体は遺伝子産物のみでできているわけではないからである。
生命活動の最も重要なエネルギー源は糖と脂質であるが、
糖と脂質は
エネルギー源以外にも生命体を構成する基材として必須である。
糖や脂質は遺伝子で定義されているのではなく、
遺伝子産物である酵素タンパク質の働きで鋳型なしに合成される。
例えると、生物は家を建てるのに家の設計図を用意するのではなく、
大工さん(=酵素タンパク質)の設計図を持っているのである。
どのような家を作るかは大工さんに任せているわけである。
このような現象は自己組織化とよばれる。
かなり大胆なことを平気でやっているが、出来てくる家(=生命体)
は殆んど同じといってよいものができてくるので不思議である。
生命の設計図(=ゲノム)には、大工さんの性能、数、
働く順序が規定されていると思われるが、
数千種類の大工さんをどのように指揮しているのであろうか。
生命体形成は、ちょうど本を書くようにいくつかの 階層 で出来ている。
文字(原子)が組み合わさって単語(分子)ができ、 単語(分子)
が組み合わさって文(分子複合体)ができ、文(分子複合体)
が組み合わさって段落(オルガネラ)ができ、 段落(オルガネラ)
が組み合わさって、節、章(細胞、組織)ができ、
それらが組み合わさって本(個体)ができる。 本の内容を理解するのに、
文字や単語のことをいくら詳しく調べてもわからない。
少なくとも文のレベルで知る必要がある。
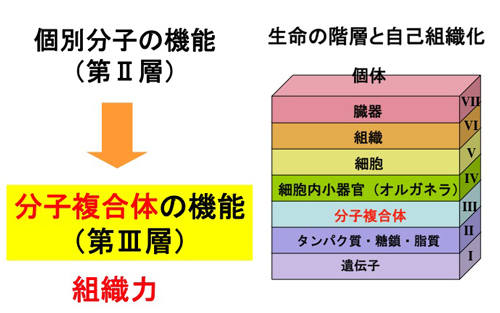
糖鎖や脂質を取り巻く状態は分子複合体の階層にあたり、
まさに、化学物質が生き物に変わる段階である。
このプロセスを科学的に実証するのに最も相応しい研究対象が生体膜である。
生体膜は、リン脂質を基本成分とする油の膜(膜脂質)から成り、そこに膜タンパク質が挿入されて出来ている。膜脂質や膜タンパク質には糖鎖が付いているものがあり、それぞれ 糖脂質、 糖タンパク質と呼ばれる。生体膜は流動的で、膜タンパク質は生体膜上をダイナミックに移動する。しかし、けっして生体膜はランダムで均一な構造体ではなく、ところどころに分子の集合体をつくる。つまり、秩序が生じる。このように分子が集まった微小領域を 膜マイクロドメインという。この膜マイクロドメインを介して膜の内外の情報伝達が行われるので、生体にとってとても重要な場である。
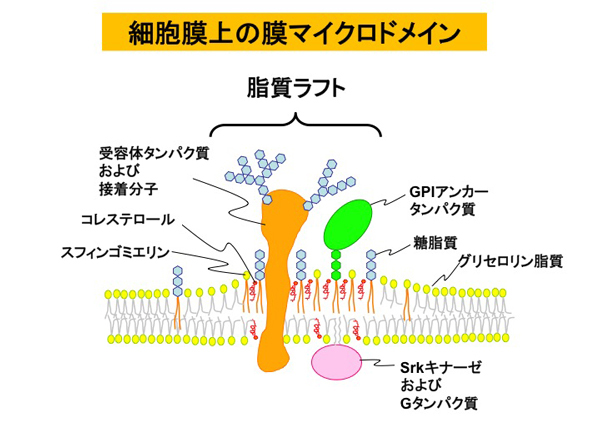
膜マイクロドメインは静的構造体ではなく、ミリ秒単位で作っては壊すを繰り返す。まさしく、生体膜は『生きている』。実際の生体において、膜マイクロドメインがどのような原理で出来るのかを研究することが生体膜研究の重要課題である。
生体膜をつくる糖鎖と脂質の異常は膜機能に変調を来し、実際、ヒトの病気をひき起こすので、これらの疾患の診断と治療を行うことは医学上も重要である。糖鎖と脂質は遺伝子産物でなく、種間を越えて共通なものも多いので、外から薬として投与することにより補填できる場合も多い。
|

