2013年12月の魚

リュウグウノツカイ Regalecus russelii (Cuvier, 1816) (アカマンボウ目リュウグウノツカイ科)
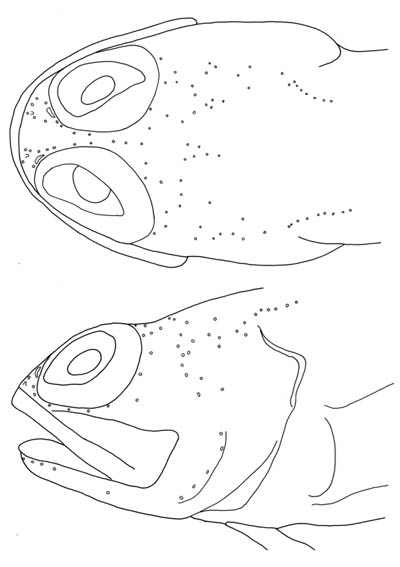
よく知られる深海魚のリュウグウノツカイは1属1種で,その学名は Regalecus glesne とされていました(Nelson, 2006).しかし,日本では "Regalecus russellii" の学名が長らく使われてきました.昨年出版されたRoberts (2012)によるリュウグウノツカイ属のモノグラフでは,これら2種は形態学的にも遺伝学的にも明瞭に異なり,それぞれ有効種であることが示されています.リュウグウノツカイは R. glesne とは次の形態形質で識別できます:腹部最後まで(肛門直上まで)の背鰭鰭条数(80以下 vs. R. glesne では 90-120),腹椎骨数(34-37 vs. 45-56),大型個体の第1鰓耙数(47-60 vs. 33-47),完全な背鰭鰭条数(333-371 vs. 414-449),背鰭最初の鰭膜でつながる伸長鰭条数とその直後の鰭膜でつながらない伸長鰭条数(3-6と1 vs. 6-8と5-11)(Roberts, 2012).また,Roberts (2012)は本属の分類の再検討の中で,本研究室の写真標本を R. russelii のネオタイプに指定しました.両種は三大洋に分布し,インド洋での記録はオーストラリア西部と南アフリカ沿岸を除いて種の同定も不明ですが,大西洋の熱帯海域,太平洋の北部から熱帯水域では R. russelii のみが,大西洋と南太平洋の高緯度海域では R. glesne のみが分布し,南アフリカ沿岸では両種の記録があります(Roberts, 2012).R. russelii のタイプ産地はインド西岸のゴア付近ですが,ネオタイプとすべきインド洋産の標本がなかったため,日本産で状態のよい本標本が指定されたわけです.
リュウグウノツカイ属は,体が極めて細長く,巨大なタチウオ型で,尾部末端が完全に残った標本がほとんどありません.しかし,全長2m以下(多くは1m以下)の若魚では,尾部骨格が観察されています (Roberts, 2012).このような尾部の欠損や採集時の“千切れやすさ”は,捕食者による損傷に適応した“自切”ではないかと以前から考えられてきました(Roberts, 2012).しかし,リュウグウノツカイの大型個体がサメ類やクジラ類などの捕食者に食べられているという報告や観察はありません.実際に,尾部の椎体や血管棘など結合が緩い骨格要素や体側筋の筋節を隔てる構造,そして“く”の字状の滑らかな欠損跡の観察からも“自切”する特異な能力が示唆されます.リュウグウノツカイ科に含まれるもう一方の属の Agrostichthys でも,同様に尾部を“自切”するようです.
リュウグウノツカイ属の2種のうち,R. glesneは表層近くから水深500m付近まで,タチウオ類のように垂直に定位した個体の観察例が知られています(Roberts, 2012).今年の6月末に沖縄県の宜野湾市で開催された第9回インド-太平洋魚類会議において,Tyson Roberts 博士は口頭発表で R. glesne が背鰭を波上に動かしながら,垂直に定位したまま上下に移動する行動のビデオ映像をクラシック音楽と共に流して喝采を浴びていました.これまでにオキアミ類を大量に食べていた個体も知られることから,表層から中深層の間を鉛直移動しているのかもしれません.生態や行動,繁殖,生活史については,依然として謎の多い魚です.
リュウグウノツカイの学名の種小名は,原記載では “russelii ”( l が1つ)と綴られ,その後多くの出版物では“russellii ”(l が2つ)が使われていました.Roberts (2012)は長い間多くの書物や論文で使用された後者の "russellii "を用いましたが,Eschmeyer (2013) のCatalog of fishes のデータベースを見ると,原綴りの “russelii ”が使われています.その理由として,「Apparently this is the first Latinization of "Russelian Gymnetrus" of Shaw 1803..."」との記述があります.Cuvier (1816)の"Gymnetrus russelii"の原記載は,Russell (1803)の図版とShaw (1803)の記載を基にしたものですが(著者が直接標本を観察していない),著者のCuvier がラテン語化した“ラッセル氏の Gymnetrus” の意味で,種小名の原綴りを "russelii"としたとの判断です.もし Cuvier がラッセル氏に献名して原綴りを誤って russelii としたならば,国際動物命名規約第4版(日本語版,2000年)の条32.5の「訂正しなければならない綴り(不正な原綴り)」となると思いますが,そういった事例ではないようなので,この原綴りが「正しい原綴り」となります.国際動物命名規約第4版の条32.5の例には,「仮に,新しい種階級群名を提唱したときに著者がその種をLinnaeusにちなんで命名すると述べたがその学名が ninnaei として公表されたとするならば,それは,linnaei と訂正すべき不正な原綴りである.」とあります.Roberts (2012)を見て“russellii ”で正しいと考えていましたが,今回調べてみたところ, “russelii ”でよいことがわかりました.崎山・瀬能 (2012)や日本産魚類検索の第3版(林,2013)では Eschmeyer (2013)に従い,russelii と正しく綴られています.
参考文献
Eschmeyer, W. N. (ed.). 2013. Catalog of fishes: genera, species, references. (http://research.calacademy.org/research/ichthyology/catalog/fishcatmain.asp). Electronic version accessed 27 Dec. 2013.
林 公義.2013.リュウグウノツカイ科.中坊徹次 (編), pp. 480, 1866. 日本産魚類検索 全種の同定. 第3版. 東海大学出版会, 秦野.
Nelson, J. S. 2006. Fishes of the world, 4th ed. Wiley, Hoboken. 601pp.
Roberts, T. R. 2012. Systematics, biology, and distribution of the species of the ocean oarfish genus Regalecus ( Teleostei, Lampridiformes, Regalecidae). Mémoires du Muséum national d'Histoire naturelle, Tome 202. Publications Scientifiques du Muséum, Paris. 268pp.
崎山直夫・瀬能 宏.2012.相模湾におけるリュウグウノツカイ(アカマンボウ目リュウグウノツカイ科)の記録について.神奈川県自然誌資料(33): 95-101. PDF
リンク Video of the Oarfish, Regalecus glesne http://www.youtube.com/watch?feature=player_embedded&v=-yIWfCAC5y0 (Youtube)
写真標本データ: BSKU 39999, 394+ cm TL, ネオタイプ,土佐湾(香南市沖,旧吉川村沖の定置網),1984年8月24日.
*沖縄での第9回インド-太平洋国際会議後,沖縄美ら海水族館でのTyson R. Roberts 博士の写真1・写真2(大水槽前),沖縄美ら海水族館の R. russelii の標本.会議後のフィールドエクスカージョンで水族館見学ツアーに参加したのですが,館内でなぜかRoberts博士に捕まってしまい,展示標本まで案内して,さらに展示標本の全背鰭鰭条を数えるはめになりました.肛門前の背鰭鰭条数は80以下で,全鰭条数は370ぐらいだったので,間違いなくリュウグウノツカイ R. russelii でした.展示標本のうち,上の大型標本は沖縄近海産,下の小型標本は高知沖産です.
(遠藤広光)
{kind=link}
{kind=link}
{kind=link}
2013年11月の魚

ヨロイワニギス Champsodon pantolepis Nemeth, 1994 (スズキ目ワニギス亜目ワニギス科)
ワニギス属 (Champsodon) はワニギス科唯一の属で, 太平洋およびインド洋の温・熱帯海域のおよそ水深 30-1100 mの大陸棚から斜面上部で見られる小型の底生性魚類です. また,夜間には表・中層に浮上することが知られています(Smith, 1953 ; Nemeth, 1994 ; Morohoshi and Sasaki, 2002). 神奈川県立生命の星・地球博物館魚類写真データベースには,2002年2月9日に静岡県駿河湾の湾内の水深3mで撮影されたワニギスChampsodon snyderi Franz, 1910の写真が掲載されています. 本属には世界で13種が知られており, そのうち日本にはワニギス, ミナミワニギス Champsodon longipinnis Matsubara and Amaoka, 1964, クロワニギス Champsodon guentheri Regan, 1908, そしてヨロイワニギスChampsodon pantolepis Nemeth, 1994 の4種が分布します (Nemeth, 1994 ; Nelson, 2006 ; 波戸岡・荻原, 2013). また,本属魚類の形態的特徴は, 体が側偏し, 斜めに裂けた大きな口と前鰓蓋棘をもち, 頭部と体の大部分が小歯状の櫛鱗で覆われ,体側には突起状の側線系が存在しています(Nemeth, 1994).
ヨロイワニギスはオーストラリア西部, 静岡県戸田, 高知県御畳瀬, 宮崎県戸々呂で採集された標本を基に記載されました(Nemeth, 1994).本種の特徴は,体側の側線系周囲も含む体全体的が鱗に覆われることです. 種小名の pantolepis の由来もギリシャ語のpantos (すべて), lepis (鱗)と体のすべてが鱗に覆われている様からつけられました(Nemeth, 1994). 本属の多くの種では腹部や側線系周辺に無鱗域をもちます.さらに,本種は頭部背側にみられる隆起骨の間に4個の感覚突起の横列とその後方には半円状に並ぶ4個の感覚突起をもつこと,胸鰭の軟条が黒色色素で覆われること, 主上顎骨の後端が眼の後縁下に達すること,そして尾椎骨の第1から第5血管棘が板状に変形することが特徴です. 本種はMatsubara et al. (1964) でクロワニギスの記載に用いられた標本にまぎれていたことから,クロワニギスによく似ており,誤同定されている場合が多々あります(Nemeth, 1994). ワニギス属のなかでも大型種で, 標準体長は100mmを超える個体も多く,今回の写真個体は167.9mm SLとこれまで知られる標本では最大かもしれません. 本種の分布は, オーストラリア西部, 台湾, 宮崎県から神奈川県まで南日本の太平洋岸沖で確認されています (Nemeth, 1994 ; 鹿児島大学総合研究博物館所蔵標本).
ワニギス属魚類は食用には向かず, 漁獲されてもゴミとして捨てられることがほとんどです.しかし,ワニギスは量的にも多く, ワニエソやタチウオ, キントキダイなどの魚食性魚の餌として重要な価値があります(山田ほか, 2007). 個人的には一度食べてみたい魚です.
参考文献
波戸岡清峰・萩原清司 2013. ワニギス科. 中坊徹次 (編), pp. 1272-1273, 2094. 日本産魚類検索 全種の同定. 第3版. 東海大学出版会, 秦野.
Matsubara, K, A. Ochiai, K. Amaoka and I. Nakamura. 1964. Revisional study of the tirachinoid fishes of the family Champsodontidae from the waters around Japan, and Tonking Bay. Bull. Misaki Mar. Biol. Inst., Kyoto Univ., 6: 1-17, pls. I-III.
Nelson, J. S. 2006. Fishes of the world, 4th ed. Wiley, Hoboken. 601pp.
Nemeth, D. 1994. Systematics and distribution of fishes of the family Champsodontidae (Teleostei: Perciformes), with descriptions of three new species. Copeia, 1994(2):347-371.
Nemeth, D. 2001. Champsodontidae: Species identification guide for fishery purposes. The living marine resources of the western central Pacific. Bony fishes part 4 (Labridae to Latimeriidae), estuarine crocodiles, sea turtles, sea snakes and marine mammals. FAO, Rome. 6: v+3497-3499.
Morohoshi, Y. and K. Sasaki, 2003. Intensive cannibalism and feeding on bregmacerotids in Champsodon snyderi (Champsodontidae): evidence forpelagic predation. Ichthyol. Res. 50: 387-390.
Smith, J. L. B. 1953. The sea fishes of southern Africa. Central News Agency, Ltd., South Africa., 175-176.
山田梅芳・時村宗春・堀川博史・中坊徹次 2007. 東シナ海・黄海の魚類誌. 東海大学出版会, 秦野. 1262pp.
写真標本データ: BSKU 94588, 167.9 mm SL, 2008年4月21日,高知市御畳瀬魚市場大手繰り網(足摺岬東方沖,水深約300m),採集&写真:中山直英.
(鰺坂晃成)
2013年10月の魚

ヘラヤガラ Aulostomus chinensis (Linnaeus, 1766) (トゲウオ目ヘラヤガラ科)
ヘラヤガラ科にはヘラヤガラ属のみが含まれ,世界で1属3種が知られています.熱帯から亜熱帯を中心にインド洋・太平洋のアフリカ東岸からアメリカ西岸まで,北は中部日本から南はオーストラリア南岸までと分布域は極めて広く,おもに造礁サンゴ類のある浅海域に生息しています (Kimura and Peristiwady, 2000; 瀬能,2013).本属3種のうち,日本にはヘラヤガラ Aulostomus chinensis (Linnaeus, 1766) のみが出現します(Kimura and Peristiwady, 2000; Bowen et al., 2001; 瀬能,2013).本種はインド洋・西太平洋に分布し,南日本,ハワイ諸島,ロードハウ島,イースター島などの島嶼の浅瀬の岩礁やサンゴ礁に生息しています (Fritzsche and Thiesfeld, 1999).
本科の体は側扁して細く長く,口は長く伸びた吻の先に位置し,斜めに開きます.下顎には1本の短い肉質の髭をもちます.長く大きな頭部は無鱗で,体は小さな櫛鱗で覆われ,側線は明瞭です(Kuiter, 1993; Nelson, 2006; 荒賀, 2007).ヘラヤガラは脊椎骨が62~64個あり,最初の4個は長い頭部を支えるために著しく大きくなり,細長く癒合しています.背鰭は2基で,第1背鰭には8~13個の短い分離棘が等間隔で並び,それぞれに三角状の鰭膜を備えます.第2背鰭には23~28本の軟条を,臀鰭には23~29本の軟条をもち,2つの鰭は尾鰭の前方で相対します.また,腹鰭は小さく,腹位にあります.背鰭と臀鰭はその基底付近を除き透明で,尾鰭には通常2個の黒斑があります.幼魚と成魚また個体間や環境により体色に変異がみられ,多くの個体では茶色で不規則な明るい縦縞をもちますが,白い横縞をいくつかもつ個体や体全体が黄色の個体もみられます.ヘラヤガラは最大で全長 80cmまで,平均では40cm程度に成長します (Fritzsche and Thiesfeld, 1999; 瀬能,2013).
ヘラヤガラは狡猾な捕食者で,餌になる小魚や甲殻類に近付くために海藻の茂みやサンゴ類の群体の間で倒立したり,他の中型から大型の魚類の背側に寄り添って餌に近付いていき,そして長い吻で一気に吸い込んで捕食します(Kuiter, 1993; Fritzsche and Thiesfeld 1999; 荒賀, 2007).写真の個体は,今年の9月24日から26日まで行われた高知県宿毛市沖の島での魚類採集に参加した研究室OBの高橋佳大さん(現在,高知大大学院農学専攻修士2年)により採集されました.本個体に近づいても逃げられることなく,簡単に手網で捕まえることができたそうです.食用魚としてのヘラヤガラは,皮膚に渋みがあるようです.定置網などで時々混獲される程度であり,商業価値はなく,地元で消費されるようです.
参考文献
荒賀忠一.2007.ヘラヤガラ科.岡村 収・尼岡邦夫 (編), p. 185. 山渓カラ―名鑑 日本の海水魚. 第三版. 山と渓谷社, 東京.
Bowen, B. W., A. L. Bass, L. A. Rocha, W. S. Grant and D. R. Rpbertson. 2001. Phylogeography of the trumpetfihes (Aulostomus): ring species complex on a global scale. Evolution, 55(5): 1029–1039.
Nelson, J. S. 2006. Fishes of the world. Forth edition. John Wiley and Sons, Inc. Hoboken, New Jersey. 601pp.
Fritzsche, R. A, and K. G. Thiesfeld. 1999. Bony fishes. Part 2: Mugillidae to Carangidae. Family Aulostomidae Vol. 4. Page 2277. In K. E. Carpenter and V. H. Niem, eds. FAO species identification guide for fisheseries purpose, the living marine resources of the Western Central Pacific. Roma, FAO.
Kuiter, R. H. 1993. Seahorses and Pipefishes order Syngnathiformes, Trumpetfishes Family Aulostomidae. The complete Divers’&Fishermen’s Gide to Coastal Fishes of South-Eastern Australia:79.
Kimura, S. and T. Peristiwady. 2000. Fishes Ikan Family Aulostomidae Pages169-170. In K. Matsuura, O. Kurnaen, Sumadhiharga and K. Tsukamoto, eds. Field guide to Lombok Island, identification guide to marine organisms in seagrass beds of Lombak Island, Indonesia.
瀬能 宏.2013.ヘラヤガラ科.中坊徹次(編),pp. 609, 1906.日本産魚類検索図鑑 全種の同定.第3版,東海大学出版会.秦野.
写真標本データ:BSKU0111673,792.5mmSL,2013年9月25日,高知県宿毛市沖の島赤崎,スクーバ潜水による手網採集,採集:高橋佳大 写真:三澤 遼.
(川内貴斗)
2013年9月の魚

リュウグウハダカ Polymetme elongata (Matsubara,1938) (ワニトカゲギス目ギンハダカ科)
ギンハダカ科ギンハダカ属魚類は深海の中深層に分布し,世界では6種が知られ,そのうち日本ではギンハダカ Polymetme corythaeola (Alcock,1898),リュウグウハダカ Polymetme elongata (Matsubara,1938),そしてタマブキイワシ Polymetme surugaensis (Matsubara,1943)の3種が報告されています.本属は囲眼部発光器(ORB)のうち,眼前発光器(PO)のみをもつことや,背鰭起部が臀鰭起部より前方にあること,腹鰭起部が背鰭起部よりも前方であることなどの点で,ギンハダカ科の他属と区別することができます(Parin and Borodulina, 1990).
リュウグウハダカは福島県沖,相模湾,駿河湾,熊野灘,土佐湾,沖縄舟状海盆,東シナ海,フィリピン諸島の中深層に生息し,遊泳生活を送ります(藍澤・土居内, 2013).本種は白味を帯びた腹膜に黒色斑が散在すること,体が細長く,体長が頭長の約5倍であることなどで,ギンハダカとタマブキイワシから区別することができます(Matsubara,1938; Parin and Borodulina, 1990).本種は熊野灘で採集された標本に基づき,京都大学の松原喜代松博士により,Yarrella blackfoldi Goode and Bean,1896の亜種として記載されました(Matsubara,1938).その後の検討により,Polymetme McCulloch,1926へと帰属が変更になり,独立種とされました。本種を含むギンハダカ科魚類は透明で大きな鱗をもちますが,とてもはがれやすいため(Matsubara 1940),水揚げされた時には鱗がほとんど残っていません.鱗のない状態では,体側上部は茶色で,下部は銀色をしています.ハダカイワシ類などと間違えられがちですが,食用とはならないので商業的価値はありません.
本種は深海のなかでも中深層(水深200mから1,000m)という,わずかに光が届く水深で生活しています.そのため,上方からの光によってできる自身の影が,下方の捕食者から見つからないように,腹部に沿った発光器を光らせることで,影を相殺しています.また,幅が狭く細長い体や銀色の体色は,少しでも影を作らないようにするためだと考えられています.このような特徴は,他の中深層に生息する魚類にも多く見られます.
参考文献
藍澤正宏・土居内龍.2013.ギンハダカ科.中坊徹次(編), pp. 381-384, 1838-1839. 日本産魚類検索全種の同定. 第3版. 東海大学出版会, 秦野.
Matsubara, K. 1938. Studies on the deep-sea fishes of Japan. VI. On some stomiatoid fishes from Kumano-Nada. Journal of the Imperial Fisheries Institute, 33 (1): 37-52.
Matsubara, K. 1940. Studies on the deep-sea fishes of Japan. XIII. On Prof. Nakazaw's collection of fishes referable to Isospondyli, Iniomi and Allotriognathi (1). Suisan Kenkiu-Shi v. 35 (12): 314-319.
McCulloch, A. R. 1926. Report on some fishes obtained by the F. I. S. "Endeavour" on the coasts of Queensland, New South Wales, Victoria, Tasmania, South and South-Western Australia. Part V. Biological Results Endeavour, 5 (pt 4): 157-216, pls. 43-56.
Parin, N. V. and O. D. Borodulina 1990. Review of the genus Polymetme (Photichthyidae) with the description of two new species. Voprosy Ikhtiologii, 30 (5): 733-743. [In Russian. English translation in Journal of Ichthyology, 30 (6):108-121.]
写真標本データ:BSKU 92885,123.5 mm SL, 高知市御畳瀬魚市場,大手繰り網,幸成丸,2007年12月17日,採集・撮影:中山直英.
(森 彩乃)
2013年8月の魚

ハネダホタルジャコ Acropoma hanedai Matsubara, 1953 (スズキ目ホタルジャコ科)
ホタルジャコ科ホタルジャコ属魚類は温帯の大陸棚周辺域に分布し,世界では5種が知られ,そのうち日本ではホタルジャコAcropoma japonicum Günther, 1859、ハネダホタルジャコ Acropoma hanedai Matsubara, 1953,オキナワホタルジャコAcropoma lecorneti Fourmanoir, 1988の3種が報告されています(Yamanoue and Matsuura, 2002; Okamoto and Ida, 2002; 波戸岡, 2013).本属は腹部に発光腺,レンズおよび反射層からなる発光器をもつことで,ホタルジャコ科の他属と区別できます(Okamoto and Ida, 2002).また,これまで本属の背鰭は第1背鰭が8棘と第2背鰭が1棘10軟条(鰭式では Ⅷ-I, 10)とされていました(例えば,山川, 1985; 山田ほか, 2007).一方,Okamoto and Ida (2002)は本属の標本を精査した結果,第1背鰭の第7棘と第8棘が鰭膜でつながらないことを明らかにし(第8棘は独立する),同属の背鰭の鰭式をⅦ-Ⅰ-Ⅰ,10としました.
ハネダホタルジャコは駿河湾から豊後水道の太平洋沿岸や沖縄舟状海盆,台湾南部,東沙群島の大陸棚縁辺から斜面域にかけての水深100~250mに生息するホタルジャコ科魚類です(波戸岡, 2013).本種は肛門が閉じた腹鰭の先端近くにあること,発光腺が長いこと,発光腺がU字型であるなどによって,ホタルジャコとオキナワホタルジャコから区別できます(山田他, 2007; 波戸岡, 2013).本種とホタルジャコは,発光腺内に共生する発光細菌により発光することが知られています(羽根田, 1895).本種のように腹部に発光器をもつ種は発光することで自分の影を隠し,下方からの敵の眼をごまかすことができるといわれています(このことをカウンターイルミネーションあるいはカウンターシェーディングと呼ばれています).本種は熊野灘で採集された標本に基づき,京都大学の松原喜代松博士により,新種として記載されました.本種の標準和名と種小名は,横須賀市自然・人文博物館の初代館長であった羽根田弥太博士に献名されたものです.羽根田博士は高知大学の前身の1つである旧制高知高等学校出身の発光生物学者で,魚類だけでなく,クラゲやホタル,キノコなど様々な生物を研究対象としました.
参考文献
羽根田弥太. 1985. 発光生物. 恒星社厚生閣. 東京. 318 pp.
波戸岡清峰. 2013. ホタルジャコ科. 中坊徹次(編), pp. 753, 1959. 日本産魚類検索全種の同定. 第3版. 東海大学出版会, 秦野.
Okamoto, M. and H. Ida. 2002. Acropoma argentistigma, a new species from the Andaman Sea,off southern Thailand (Perciformes: Acropomatidae). Ichthyol. Res., 49: 281-285.
山田梅芳・時村宗春・堀川博史・中坊徹次. 2007. 東シナ海・黄海の魚類誌. 東海大学出版会, 東京. lxxiii+1263pp.
山川武. 1985. ホタルジャコ・ハネダホタルジャコ. 岡村収(編), p. 455.沖縄舟状海盆及び周辺海域の魚類.日本水産資源保護協会,東京.
Yamanoue Y. and K. Matsuura. 2002. A new species of the genus Acropoma (Perciformes: Acropomatidae) from the Philippines. Ichthyol Res., 49: 21–24.
写真標本データ:BSKU 87802 (M5417),103.4 mm SL,2004年1月14日,高知市御畳瀬魚市場(大手繰り網).
(内藤大河)
2013年7月の魚

アカアマダイ Branchiostegus japonicus (Houttuyn,1782)(スズキ目アマダイ科)
アカアマダイはアマダイ科(Branchiostegidae)アマダイ属(Branchiostegus Rafinesque, 1815 )に属する底生性魚類です.アマダイ属の分類は研究者により見解が異なり,アマダイ科(Dooley ,1978, 1999)あるいはキツネアマダイ科 (Malacanthidae) のアマダイ亜科 (Branchiosteginae or Latilinae) とする場合があります (Nelson, 2006) .また,Imamura (2000) はキツネアマダイ科の浮遊期の仔稚魚の形態が,セミホウボウ科のそれと類似していることに着目し,比較解剖学的研究により見出した8個の共有派生形質にもとづいて,セミホウボウ科とキツネアマダイ科が姉妹群関係にあるとしました.そして,セミホウボウ科のなかにアマダイ亜科,キツネアマダイ亜科,オキナワサンゴアマダイ亜科,セミホウボウ亜科の4亜科を含め,アマダイ属はLopholatilus属とともにアマダイ亜科に分類されました.しかし,最近の遺伝子解析の結果によれば,アマダイ科アマダイ属とすることが妥当と報告されています(Dooley and Iwatsuki, 2012).
アマダイ科はCaulolatilus Gill, 1862, Lopholatilus Goode and Bean, 1880, そしてアマダイ属の3属30種からなります.アマダイ属は東部大西洋と日本を含むインド・西部太平洋の水深60-600 mの大陸棚から斜面上部に広く分布し,現在17種が知られています.他の2属は西部大西洋と東部太平洋にのみ分布します(Dooley, 1978, 1998; Nelson, 2006; Dooley and Iwatsuki, 2012; Hiramatsu and Yoshino, 2012).アマダイ属魚類は,体が延長する四方形であり,明瞭な頭部背中線をもつこと,背鰭条数が22(6–8棘,14–16軟条),臀鰭条数は14(1–2棘,12–13軟条),有孔側線麟数は45-72枚,脊椎骨数は10+14,尾鰭は載形か二重載形である特徴をもちます(Dooley, 1978; 平松, 2002).
アカアマダイは南シナ海から日本まで広く分布し,東アジアの水深60-200 mの大陸棚に生息しています.日本周辺では,日本海側は青森まで,太平洋側は千葉県までの海域に分布します(Dooley, 1978; 堀川ほか,2001;Chen et. al. 2002; 藍澤・土居内,2013).現在,日本周辺海域に分布するアマダイ属には,アカアマダイの他に,スミツキアマダイ B. argentatus (Cuvier in Cuvier and Valenciennes, 1830),キアマダイ B. auratus (Kishinouye, 1907),シロアマダイB. albus Dooley, 1978, ハナアマダイ B. okinawaensis Hiramatsu and Yoshino, 2012,の5種が知られています (Dooley, 1978, 1998; Hiramatsu and Yoshino, 2012; 藍澤・土居内,2013).アカアマダイはこれら同属他種より,頬の鱗が厚く皮膚に覆われ,その枚数は8-10枚であること,眼窩後縁に三角形の銀白色の斑紋があり,体中央に大きな黄斑をもち,尾鰭の上葉は朱色で,下葉は暗い灰白色,中央に3本の黄色縦帯をもつことで区別できます.
本種は1782年にMartius Houttuyn(ハウッタイン)によりCorypheana japonicaとして新種記載されました (Dooley, 1978).この記載に使用された標本は,1775年8月から翌年の12月まで長崎出島に医師として赴任していたThunberg(ツンベリー)により,南日本で採集されたものです.それらの標本は当時バタヴィアにいた外交官であるRadermacherを経由して,医者で博物学者のハウッタインに送られました.その時,彼は42種の動物を記載し,そのうち魚類は22新種を含む36種でした(山口・町田,2003).ハウッタインの記載は簡単で,標徴が明示されておらず曖昧ですが,現在も16種が有効種と認められており,そのうちの1種がアカアマダイです.
アカアマダイは水産上の重要種であり,白身で美味な魚として知られています(堀川ほか,2001).とくに,京都ではハモと同様に古くから珍重され,若狭の甘鯛として好まれています[京都通(京都観光,京都検定):百科事典,赤甘鯛 ぐじ)].また,静岡県中部地方でも,一夜干しにしたものを興津鯛(季語;冬)と呼称し,古くから食べてられていたようです.この“興津鯛”という地方名の由来は,甲子夜話によれば,徳川家康が献上されたアマダイ属の干物をいたく気に入り,それを献上したのが奥女中の“興津の局”であったので,その名前に由来すると言われています(日本の旬,魚のお話,甘鯛).また,別の説では,慶長年間(1596-1615年)に広瀬又右衛門がアマダイの干物を作り,それを興津河内守が家康に献上したところたいそう美味であると喜ばれ,それで興津鯛と呼ぶようになったとも言われています.どちらが正しいかはさだかでないにしろ,あの戦国の英雄の舌をも満足させた魚であったようです.さらに,江戸時代の著名な絵師である伊藤若冲は“動植綵絵”で,アカアマダイの特徴を捉えた見事な彩色画を描いています.そのほかに,曲亭馬琴もその著書,“馬琴道中記”のなかで,“このあたりもみじめずらし興津鯛“と俳句で詠んでいます.このようにアマダイ類は古くから日本人に好まれていた魚であることをうかがい知ることができます.
{kind=link}
参考文献
藍澤正宏・土居内龍.2013.アマダイ科.中坊徹次(編),pp. 867-868, 1987.日本産魚類検索図鑑 全種の同定.第3版,東海大学出版会.秦野.
Chen, G. Z. Chen, Y. Cai and S. Lin. 2002. Fishes from Nansha Islands to South China coastal waters II, Science Press, Peking. 114pp.
Dooley, J. K. 1978. Systematics and biology of the tilefishes (Perciformes: Branchiostegidae and Malacanthidae) with the description of two new species. NOAA Technical Report, NMFS circular, 411, i+v+1-78.
Dooley, J. K. 1998. Bony fishes. Part2: Mugillidae to Carangidae. Family Malacanthidae Vol. 4. Pages 2630-2648. In Carpenter, K. E. and Niem, V.H. eds. FAO species identification guide for fisheseries purpose, the living marine resources of the Western Central Pacific. Roma, FAO.
Dooley, J. K. and Y. Iwatsuki. 2012. A new species of deepwater tilefish (Percoidea: Branchiostegidae) from the Philippines, with the brief discussion of the status of tilefish systematics. Zootaxa, 3249: 31-38.
堀川博史・ Li, C.・山田梅芳・Zheng, Y.・時村宗春.2001. アカアマダイの生物学的,生態学的な比較.堀川博史,鄭元甲, 孟田湘(編),pp. 65-76.東シナ海・黄海主要資源の生物,生態特性-日中間の知見の比較.西海区水産研究所,長崎.
平松 亘.2002.アマダイ科アマダイ属魚類の分類学的再検討.高知大学大学院理学研究科生物学専攻修士論文.133pp.
Hiramatsu, W. and T. Yoshino. 2012. A new tilefish, Branchiostegus okinawaensis (Perciformes: Branchiostegidae) from Okinawa Island, southern Japan. Bull. Natl. Mus. Natr. Sci., ser A (supl. 6): 41-47.
Imamura, H. 2000. An alternative hypothesis of the phylogenetic position of the family Dactylopteridae (Pices: Teleostei), with a proposed new classification. Icthyological Res., 47 (3): 203-222.
Nelson, J. S. 2006. Fishes of the world. 4th edition. John, Wiley & Sons, Inc. Hobokon, New Jersey, 601 pp.
山口隆男・町田吉彦. 2003. Fish specimen collected in Japan by Ph. F. von. Siebolt and H. Burger and now hold by the Natural Natuurhistorisch Museum, in Leiden and other two Museum. Calanus, spec. (4): 87-337.
写真標本データ: BSKU 106454, ca. 300 mm SL, 2011年12月14日,高知市御畳瀬魚市場(幸成丸,大手繰り網),写真撮影: 中山直英
(平松 亘)
2013年6月の魚

マダラテンジクダイ Apogonichthyoides umbratilis Fraser and Allen, 2010 (スズキ目テンジクダイ科)
テンジクダイ科は世界の3大洋の温暖な海域に分布し,インド洋−太平洋にすむ魚類の中では最も大きなグループの一つです(Allen and Erdmann, 2012).本科には沿岸域の岩礁やサンゴ礁で暮らす仲間が多く,現在31属343種が記載されています(Froese and Pauly, 2013).また,この数10年で多くの新種が記載され,今後も新種記載が予想される科です.Fraser (1972) はテンジクダイ科を3亜科19属14亜属に分類し,そのうち10亜属をテンジクダイ属に含めました.その後,これらの亜属を含め多くが属に昇格しています.さらに,Pseudamia, Cheilodipterus, Archamia, Nectamia, Siphamia, そしてFoa の各属については,分類学的に再検討されました(Allen and Erdmann, 2012).日本においては2亜科18属93種(うち1未記載種を含む)が報告されています(林, 2013).
Fraser and Allen (2010)はApogonichthyoides Smith, 1968を復活させて,本属内に2種群19種を認めました.その後,本属は22種となり,これまでにApogonとされたクロイシモチ(A. niger)も本属に移されています(Fraser, 2010; Fraser and Allen, 2011).マダラテンジクダイは,Fraser and Allen (2010)により西オーストラリアのノースウエスト礁を基産地(ホロタイプの産地)として,Apogonichthyoidesの新種として記載されました.Yamakawa and Manabe (1986)は,高知県柏島で採集された標本(KSHS20353)をカクレイシモチApogon fraxineusとして報告しましたが,この標本は本種と判明しました(林,2013).また,Fraser and Allen (2010)は Apogon timorensis (カクレテンジクダイ)を本属に移し,その後林(2013)は本属にカクレテンジクダイ属の新称を与え,日本から本種とカクレテンジクダイの2種を報告しています.カクレテンジクダイ属は尾鰭が浅い湾入形で,第2背鰭と臀鰭には黒色斑がなく,頬部には暗色斜帯があることで,他の属と見分けられます.日本に分布する2種は,マダラテンジクダイでは胸鰭の鰭条数が14であること(カクレテンジクダイでは16),体側と尾柄部にそれぞれ2~3本の不明瞭な暗色横帯があること(カクレテンジクダイではどちらも2本の明瞭な横帯),頬部の暗色斜帯が不明瞭(カクレテンジクダイでは明瞭な1本)などで識別できます.
日本でのマダラテンジクダイの記録は,四国西南部沿岸の2箇所のみで,高知県大月町柏島では,カクレイシモチ Apogon fraxineus として報告された4標本が採集され(40.0−55.0 mm SL;Yamakawa and Manabe, 1986),愛媛県愛南町ではテンジクダイ科の一種(未同定属および種)として水中写真が撮影されました(高木ほか,2010).海外ではパラオ諸島,インドネシア,ブルネイ,ラジャアンパット島(ニューギニア島西部),オーストラリア西岸からの報告があります(Allen and Erdmann, 2012).海外では,水深10~40mにおいて,単独もしくはペアで泳ぐ姿が観察されていますが,日中は岩や礫の間に隠れています.標本は転石や死んだ造礁サンゴのがれ場,ハードコーラルの少ない,海綿やヤギ類が豊富な沿岸から数キロの離礁で採集されています.また,それらの中にはハナガササンゴ類やクサビライシ類,大型藻類の豊富な平らな砂地や貝殻の下などから採集されたものもあります(Fraser and Allen, 2010).
写真標本が採集された柏島でも,本種は水深 8~10 mで,単独もしくはペアで生息していますが,日中には死んだ造礁サンゴ類のがれ場,転石や砂地の廃棄物の中に隠れており,観察の頻度は非常に少ないです.一方,夜間には砂地に現れ,比較的普通に観察することができます.愛南町の報告と同様に,柏島でも口内保育中の個体は6~7月に,幼魚は8~10月頃に観察されています.本種はインド−西太平洋の分布域が広いことから,今後は日本の他地域からの報告例も増えることが予想されます.
参考文献
Allen, G. R. and M. V. Erdmann
. 2012. Reef fishes of the East Indies. Vol. I-III. Tropical Reef Research, Perth. x+1-424pp. +end note, 425-855pp. preface, map, contents and 857-1260pp.
Fraser, T. H. 2010. A new deepwater species of the genus Apogonichthyoides (Apogonidae) from the Timor Sea, northern-western Australia. Aqua, Int. J. Ichthyol., 16(3): 93–100.
Fraser, T. H. and G. R. Allen. 2010. Cardinalfish of the genus Apogonichthyoides Smith, 1949 (Apogonidae) with a description of a new species from the West-Pacific region. Zootaxa, 2348: 40-56.
Fraser, T. H. and G. R. Allen. 2011. A new cardinalfish of the genus Apogonichthyoides (Perciformes, Apogonidae) from Raja Ampt Islands, with a key to species. Zootaxa, 3095: 63-68.
Froese, R. and D. Pauly, eds. 2013. FishBase. World Wide Web electronic publication: www.fishbase.org, version (04/2013).
林 公義. 2013. テンジクダイ科. 中坊徹次 (編), pp. 826-864, 1979-1986. 日本産魚類検索 全種の同定. 第3版. 東海大学出版会, 秦野.
高木基裕・平田智法・平田しおり・中田 親.2010.えひめ愛南お魚図鑑.創風社出版,松山.249pp.
Yamakawa, T. and S. Manabe. 1986. Forty-two rare fishes from the fish fauna of Kochi Prefecture, Japan. Mem. Fac. Sci. Kochi Univ., 7 (ser. D):
43-50, 2 pls.
写真標本データ: BSKU 110314, 30.0 mm SL, 2013年5月31日,高知県幡多郡大月町,水深 8 m,写真撮影: 松野靖子 黒バックの写真
(松野靖子・遠藤広光)
{kind=link}
2013年5月の魚

ヤイトアゴアマダイ Opistognathus trimaculatus Hiramatsu and Endo, 2013(スズキ目アゴアマダイ科)
ヤイトアゴアマダイは土佐湾の水深150–250 mから底びき網により採集された2標本を基に新種記載されたアゴアマダイ属の魚類です(Hiramatsu and Endo, 2013).アゴアマダイ科魚類はLonchopisthus Gill, 1862, アゴアマダイ属 Opistognathus Cuvier, 1816 とカエルアマダイ属 Stalix Jordan and Snyder, 1902 の3属83種と多くの未記載種を含み,東部大西洋, 地中海と中央太平洋を除く全世界の熱帯から亜熱帯海域に広く分布する底生性魚類です(Smith-Vaniz, 2010, 2011; Nelson, 2006; Allen and Erdmann, 2012).そのうち,日本にはアゴアマダイ属9種(うち3種は未記載種)とカエルアマダイ属3種の2属12種が分布します(林・大栗, 2007;藍澤・土居内, 2013).これら2属は背鰭棘の先端が針状(アゴアマダイ属)かY字状に二叉する(カエルアマダイ属)ことで簡単に区別できます (藍澤・土居内,2013).また,巣孔の形状でも両属は異なり,アゴアマダイ属は浅い垂直な孔とそこから横に分枝する複雑な孔をもちます.一方,カエルアマダイ属はほぼ垂直の深い孔を掘り,その末端がやや太くなることで異なります.さらに,アゴアマダイ属は砂礫とウニの殻や貝の破片の混じる粗めの底質を好みますが,カエルアマダイ属は細かな砂質域をおもな生息場所とする傾向があります(平松,未発表).
本種の属するアゴアマダイ属は西部大西洋,東部太平洋,インド西部太平洋の熱帯から亜熱帯の水深0.3–375 m の海域に広く分布し,本科の中で最も多様性の高い属であり,68の有効種と34の未記載種が知られています(Smith-Vaniz, 1997; Hiramatsu and Endo, 2013).本属の魚類は大きな口をもち,頭部は鱗に覆われず,背鰭は9–12棘12–22軟条,臀鰭は2–3棘10–21軟条,側線は背鰭基部に近接して後方に伸びる特徴をもちます(Nelson, 2006).また,本属の魚類はサンゴ礁域のダイバーが潜水できる範囲に生息する種が多く,海底に自分の口で垂直な穴を掘り,そこで口内保育をするという独特の生態をもつので,アマチュアのダイバーだけでなく,水中の生態写真家にとっての格好の被写体として,多くの生態写真が撮影されている魚類のひとつでもあります.
現在,日本周辺海域から知られているアゴアマダイ属はニジアマダイO. evermani (Jordan & Snyder, 1902), アゴアマダイO. hopkinsi (Jordan and Snyder, 1902), ニラミアマダイO. iyonis Jordan and Thompson, 1913, ワニアマダイ O. castelnaui Bleeker, 1860, ホソミアゴアマダイO. sp. 1,ムシクイアゴアマダイ O. sp.2, イレズミアゴアマダイ O. decorus Smith-Vaniz and Yoshino, 1985, セトアマダイ O. liturus Smith-Vaniz and Yoshino, 1985, メガネアゴアマダイO. sp. 3の9種であり,本種は日本からの10番目の種となります(林・大栗,2007;藍澤・土居内,2013).
ヤイトアゴアマダイは,背鰭が11−12棘10−11軟条,臀鰭が2棘10軟条,頭部,後頭部,胸部,胸鰭基部,背鰭の前方と側線上部が無鱗で,側線鱗数が48−50,側線の末端が背鰭第10棘と第1軟条の間にあり,上顎の後端が短く,長く葉状にならず,前鰓蓋骨に達せず,頭部の感覚孔が少なく,側線感覚孔も単孔(感覚孔の図)からなる特徴をもつことで,同属他種から識別可能です.また,本種が薄い茶色の体色にやや濃い褐色の5本の横帯と背鰭基底に3個の暗色斑をもつことも,他種との識別点です.本種はニジアマダイ,セトアマダイ,O. hongkongiensis Chan, 1968, O. rufilineatus Smith-Vaniz and Allen, 2007, O. crassus Smith-Vaniz, 2010の5種と同数の背鰭と臀鰭の鰭条数をもちますが,ニジアマダイとO. rufiliniatusとは,体側に5本の暗色横帯があること,背鰭の基部に3個の暗色斑があることで区別できます.また,O. hongkongiensis とO. crassus は本種と同様に体側に横帯と背鰭に暗色斑をもちますが, 前者は横帯の数が7本をもつことで,後者は背鰭基部に5個の暗色斑をもち,頭部の感覚孔の数が多いことで異なります.さらに,セトアマダイとは頭部に暗色斑をもたないことで区別できます.土佐湾や柏島周辺の海域から未記載種と考えられるアゴアマダイ属の魚が採集され,また生態写真も撮影されているので,今後も分類学的研究の必要な魚類の一つです.
{kind=link}
参考文献
藍澤正宏・土居内龍. 2013. アゴアマダイ科. 中坊徹次 (編), pp. 815-818, 1976. 日本産魚類検索 全種の同定. 第3版. 東海大学出版会, 秦野.
Allen, G. R. and M. V. Erdmann
. 2012. Reef fishes of the East Indies. Vol. I-III. Tropical Reef Research, Perth. x+1-424pp. +end note, 425-855pp. preface, map, contents and 857-1260pp.
Chan, W. L. 1966. Note on opistognathid jawfishes from Hongkong, with the description of a new species. Japanese Journal of Icthyology, 14(1/3): 9-11.
林公義・大栗智史.2007.日本産アゴアマダイ属(アゴアマダイ科)魚類の分類 学的再検討.横須賀市博物館研究報告(自 然),54:27-57.
Hiramatsu, W. and H. Endo. 2013. Opistognathus trimaculatus, a new jawfish (Teleostei: Opistognath- idae) from Tosa Bay, Japan. Bulletin of the National Museum of Nature and Science, Ser. A, (Suppl. 7): 19-23.
Nelson, J. S. 2006. Fishes of the world. 4th edition. John Wiley & Sons, Inc. Hoboken, New Jersey. 601 pp.
Smith-Vaniz, W. F. 1997. Five new species of jawfishes (Opistognathus: Opistognathidae) from the western Atlantic Ocean. Bulletin of Marine Science, 60 (3): 1074-1128.
Smith-Vaniz, W. F. 2010. New species of Indo-Pacific jawfish (Opistognathidae) from the Western Indian Ocean and Red Sea. Smithiana Bulletin, (12): 39-54.
Smith-Vaniz, W. F. 2011. Opitognathus albicaudalis, a new jawfish (Teleostei: Opistognathidae) from Andaman Islands. Zootaxa, 3085: 34-40.
Smith-Vaniz, W. F. and G. R. Allen. 2007. Opistognathus ruflineatus, a new species of jawfish (Opistognathidae) from the Bird's Head Peninsula, western New Guinea. Aqua International Journal of Ichthyology, 13 (1): 18-27.
写真標本 :NSMT-P 111154 (BSKU 41806を移管),72.0 mm SL,ホロタイプ,土佐湾中央部,調査船こたか丸,オッタートロール,水深約150 m,1985年5月23日.
(平松 亘)
2013年4月の魚

アオザメ Isurus oxyrinchus Rafinesque, 1810 (ネズミザメ目ネズミザメ科)
ネズミザメ目 (Order Lamniformes) には世界で7科10属15種が,日本周辺からはそのうち7科10 属13種が知られています.7科のうち4科はそれぞれ1属1種からなり,深海性のミツクリザメMitsukurina owstoni Jordan, 1898,巨大な濾過食者であるメガマウスザメMegachasma pelagios Taylor, Compagno and Struhsaker, 1983やウバザメCetorhinus maximus (Gunnerus, 1756),そしてミズワニ Pseudocarcharias kamoharai (Matsubara, 1936),です.残る3科は数種を含むネズミザメ科,オナガザメ科やオオワニザメ科であり,ネズミザメ目は多様な形態や生態を見せる種から構成されています(Nelson, 2006).
ネズミザメ科 (Family Lamnidae)は人食いザメとして有名なホホジロザメCarcharodon carcharias (Linnaeus, 1758) を含み,我々人間にとって比較的危険なグループです (仲谷,2011).ネズミザメ科は日本ではホホジロザメ,ネズミザメLamna ditropis Hubbs and Follet, 1947,アオザメ,バケアオザメIsurus paucus Guitart Manday, 1966の3属4種が知られ(アオザメ属は本種とバケアオザメのみ),バケアオザメを除く3種は日本海側および太平洋側の北海道周辺から九州まで広く分布しています.アオザメはさらに琉球列島まで出現します.このグループは遊泳性が強く,沿岸域から外洋の表層付近から水深650 mまで出現し,世界の熱帯から温帯域まで広く分布しています (吉野ほか,2013). アオザメの和名は特徴的な青みがかった体色に由来し,英名の“Mako”はニュージーランドのマオリ族の言葉で“サメ”を意味します.全世界でアオザメに襲われた事件は1580年から2008年まで13件あり,そのうちの1件は死亡事故でした (仲谷,2011).いっぽう,ホホジロザメによる事故は244件あり (そのうち死亡事故65件),全サメ類の中では群を抜いて危険ですが,アオザメにも注意が必要です.
アオザメの両顎歯には鋸歯縁がなく,尾柄部側面の隆起線が1本であることでホホジロザメやネズミザメと区別することができます.また,アオザメは胸鰭が短いので,胸鰭が非常に長いバケアオザメとの識別は容易です.本種の最大全長は約4 m,遊泳速度はサメの中で最も速いとされ,瞬間的には時速35 kmを越える非常に活発な種です (仲谷,2011).そのため,ゲームフィッシングの対象ともされるようです (Compagno et al., 2005).
写真の個体は今年の3月28日,研究室の先輩である朝岡さんが室戸市の鮮魚店「浦戸屋」において600円で購入しました.全長は約80 cmほどで,アオザメは全長60–70 cmで親魚から産出されるので,この個体はまだまだ子供ということになります.このように手ごろなサイズの標本は,置き場や管理に手間がかからず非常に貴重です.
鮮魚店で販売していることからも分かる通り,アオザメは食用になり,練り製品の材料やフカヒレが有名です.広島県では“ワニ料理”として親しまれており,刺身や湯引き,ワニバーガーというメニューもあるようです.私も茹でた皮をポン酢で食べてみましたが,軟骨魚類特有のアンモニア臭さはなく,寒天質に富んだ独特な食感で濃厚な味わいでした.
参考文献
吉野哲夫・青沼佳方・柳下直己・山口敦子. 2013. ネズミザメ科. 中坊徹次 (編), pp. 160, 1759. 日本産魚類検索 全種の同定. 第3版. 東海大学出版会, 秦野.
Compagno, L. J. V., M. Dando and S. Fowler. 2005. Sharks of the world. Princeton University Press, Princeton and Oxford. 368 pp.
仲谷一宏. 2011. サメ―海の王者たち―. ブックマン社, 東京. 240 pp.
Nelson, J. S. 2006. Fishes of the world. 4th edition. John Wiley & Sons, Inc. Hoboken, New Jersey. 601 pp.
パーカー,S.2010. The encyclopedia of sharks 世界サメ図鑑(仲谷一宏,日本語監修) .ネコ・パブリッシング,東京.224 pp.
写真標本 :BSKU 50684,775 mm TL,室戸市,鮮魚店浦戸屋にて購入,2013年3月28日,採集者:朝岡 隆,写真撮影:中山直英,朝岡 隆.
(三澤 遼)
2013年3月の魚

ツマグロミミズアナゴ Scolecenchelys fuscapenis McCosker, Ide and Endo, 2012 (ウナギ目ウミヘビ科ニンギョウアナゴ亜科)
2013年2月に刊行された日本産魚類検索第3版のウミヘビ科(波戸岡,2013)では,Castle and McCosker (1999)の見解に従い,旧来の日本産の“ミミズアナゴ属 Muraenichhtys Bleeker, 1853 ”の種は2分されました.そして和名がミミズアナゴのS. gymnota を含む Scolecenchelys Ogilby, 1897 に対してミミズアナゴ属が用いられ, Muraenichthys にはミナミミミズアナゴ属の和名が新たに付けられました.ミミズアナゴ属には世界で21有効種が含まれ,そのうち日本には最近新種記載された2種を含め,次の6種が分布します(McCosker et al., 2012; Hibino et al., 2012, 2013; Hatooka, 2013):ニホンミミズアナゴ S. japonicus (Machida and Ohta, 1993), クリミミズアナゴ S. macroptera (Bleeker, 1857), ミサキウナギ S. aoki (Jordan and Snyder, 1901), ミミズアナゴ S. gymnota (Bleeker, 1857), ツマグロミミズアナゴ S. fuscapenis Mccosker, Ide and Endo, 2012, そして ノドグロミミズアナゴ S. fuscogularis Hibino, Kai and Kimura, 2013.とくに,日本周辺のニンギョウアナゴ亜科(Myrophinae)の分類は,最近三重大学の研究グループによりシノニム関係の整理,再記載や新種記載が行われ,次々と論文が出版されています(例えば,Hoshino et al., 2011; Hibino et al., 2012, 2013).
ツマグロミミズアナゴは,2012年3月末に出版された国立科学博物館の新種記載プロジェクト論文集の魚類の第4弾で記載したウミヘビ科の3新種のうちのひとつです(McCosker et al., 2012).本種は水産総合研究センターの調査船こたか丸により土佐湾の水深182-269 m から採集された20標本(全長236–367 mm)と,米国スミソニアン自然史博物館に所蔵され1906年にアルバトロス号により新潟沖の水深 112 m から採集された1標本(USNM 151440, 全長 208 mm)に基づいています.本種は比較的細長く(脊椎骨数が157-162と多い),歯の特徴(2−3歯列),背鰭始部が肛門よりも前に位置し,尾部末端に特徴的な黒色斑紋をもつことから,日本産の本属他種とは容易に識別できます.学名の種小名“fuscapenis”は“黒い尾”の意味で,標準和名の“ツマグロ”も尾部後端の黒斑に因んでいます.
この論文では,ウミヘビ属のマチダウミヘビ Ophichthhus machidai と シシガシラウミヘビ O. obtusus も記載し,いずれも土佐湾沿岸で採集された標本をタイプに含んでいます.これらの3新種は,2004年3月に本研究室で大学院の修士課程を修了した井手幸子さんの「土佐湾産ウミヘビ科魚類の分類学的研究」により未記載種と判明したものですが,その後論文化されていませんでした.当時の指導教官であった町田吉彦先生(高知大学名誉教授)が退職された後,これらの種を何とか命名したいと考えていたところ,2010年11月にウミヘビ科の分類の大御所である John E. McCosker 博士(米国カリフォルニア科学アカデミー)が本研究室を訪れた際に応援を頼んで,共著で論文を出版できました.新種を発見した研究から8年後に学名と標準和名が付いたわけです.また,日本産魚類検索第3版の種の検索ページへの掲載には間に合いませんでしたが,分類学的付記の文献リストには含められています.
参考文献
Castle, P. H. J. and J. E. McCosker. 1999. A new genus and two new species of Myrophine worm-eels with comments on Muraenichhtys and Scolecenchelys (Anguilliformes: Ophichthidae).
波戸岡清峰.2013.ウミヘビ科.中坊徹次(編),pp. 266–277, 1794–1802. 日本産魚類検索全種の同定.第3版.東海大学出版会,秦野.
Hibino, Y., S. Kimura, K. Hoshino,
K. Hatooka and J.E. McCosker. 2012. Validity of Scolecenchelys aoki, with a redescription of Scolecenchelys gymnota (Anguilliformes: Ophichthidae). Ichthyol. Res., 59: 179-188.
Hibino,
Y., Y. Kai and S. Kimura. 2013. Scolecenchelys fuscogularis (Anguilliformes: Ophichthidae, Myriphinae), a new worm eel from Japan. Ichthyol. Res., 60: 43-47.
Hoshino, K., Y. Hibino, S. Kimura and Y. Machida. 2011. The worm eel, Muraenichthys okamurai Machida and Ohta 1996, a junior synonym of Muraenichhtys borealis Machida and Shiogaki 1990. Ichhtyol. Res., 58: 184-187.
McCosker, J. E. , S. Ide and H. Endo. 2012. Three new species of ophichthid eels (Anguilliformes: Ophichthidae) from Japan. Bull. Natl. Mus. Nat. Sci., Ser. A (Zool.) (Suppl. 6): 1–16.
写真標本: NSMT-P 106570 (BSKU 96046を移管), 337 mm TL, 雌,2008年8月26日,土佐湾中央部,水深223–269m,こたか丸,採集者:梨田一也・中山直英,写真:中山直英.
(遠藤広光)