2004年 12月
サンゴイワシ Neoscopelus
microchir Matsubara
(ハダカイワシ目ソトオリイワシ科) ソトオリイワシ科は,世界で3属6種,日本では2属4種が報告され,背鰭基底部が臀鰭基底部よりも著しく前にあること,上顎が前上顎骨のみで縁取られること,主上顎骨の後端が著しく幅広くなること,上主上顎骨があること,脂鰭があることなどの特徴があります.ソトオリイワシ属は,腹中線上に1列と体側に横方向に走る数列の発光器があること,舌の縁辺部に1列の発光器があることなどで,同じ科に含まれるクロゴイワシ属と区別されます.
ソトオリイワシ科は,世界で3属6種,日本では2属4種が報告され,背鰭基底部が臀鰭基底部よりも著しく前にあること,上顎が前上顎骨のみで縁取られること,主上顎骨の後端が著しく幅広くなること,上主上顎骨があること,脂鰭があることなどの特徴があります.ソトオリイワシ属は,腹中線上に1列と体側に横方向に走る数列の発光器があること,舌の縁辺部に1列の発光器があることなどで,同じ科に含まれるクロゴイワシ属と区別されます.
サンゴイワシは,太平洋,インド洋,大西洋の暖海域に広く分布し,水深300~1000 mの陸棚縁辺海域などの深海底近くに生息します.日本では駿河湾から東シナ海に分布しています.本種は,Matsubara(1943)により,1939年11月24-25日に駿河湾戸田沖で採集された標準体長146.8 mm の個体をもとに新種記載されました.本種は体側発光器が腹鰭より後ろでは1列となり臀鰭後方に達すること,胸鰭先端が背鰭基底後端に達しないことなどの特徴で他種と区別できます.本種は,陸棚縁辺直上に生息するため,しばしば深海底曳網で漁獲されます(川口,1998).本標本も土佐湾で底曳き網漁を行っている御畳瀬漁港で採集されたものです.従って,同目のハダカイワシ科魚類のように中層で生活し,長い距離を日周鉛直移動する種ではないと考えられます.本種は陸棚斜面に沿って海底と中深層を鉛直移動し摂餌するため,自分の影を消して身を守るための発光器を備えたと考えられます.ハダカイワシ科魚類は生物量が多く,大量に漁獲されるため,高知県では馴染みの“ヤケドの干物”として食用にされています.一方,サンゴイワシ科は生物量が少ないためか,多く漁獲されないので,なかなかお目に掛かれません.でも味はいけるそうです.ヤケドと同様に干物にすることをお勧めします.
参考文献
川口弘一.1998.ハダカイワシ目.中坊徹次・望月賢二(編),pp.55-57.日本動物大百科.平凡社,東京.
益田一・尼岡邦夫・荒賀忠一・上野輝彌・吉野哲夫(編).1984.日本産魚類大図鑑.東海大学出版会,東京.xx+448 pp., 370 pls.
Matsubara,
K. 1943. Ichthyological annotations
from the depth of the Sea of Japan. II. A review of the scopelid
fish, referable to the genus Neoscopelus. J. Sigenkagaku
Kenkyusyo, 1 (1) : 55-63.
Nakabo, T.
2002. Fishes of Japan with
pictorial keys to the species English edition. Tokai University
Press, Tokyo. 1xi+1749 pp.
写真個体:BSKU 73857,147.7mm
SL,2004年12月1日,高知市御畳瀬魚市場(大手繰),採集者:滝上耕太・三宅崇智・片山英里.
(三宅崇智)
2004年 11月
ムネエソ Sternoptyx
diaphana Hermann
(ワニトカゲギス目ムネエソ科) ムネエソは,ワニトカゲギス目ムネエソ科に属する全長5cm程の中深層性魚類です.本種は世界の熱帯から温帯水域にかけて広く分布し,水深
200 mから3,000 m に生息します.日本では北海道から沖縄にかけての太平洋岸沖に出現します.本科はキュウリエソ亜科とムネエソ亜科に分類され,日本周辺からは6属17種が報告されています.ムネエソ亜科は,眼が大きいこと,体が小判状で高く短いこと,体側下方に発光器が並ぶことで他の本目魚類との識別は容易です.本亜科の中でも,ムネエソ属
Sternoptyx は,他のテンガンムネエソ属 Argyropelecus やホウネンエソ属
Polyipnus に比べると,体全体が斜め後方へ傾いたように変形し(四角形から平行四辺形に),臀鰭基底部に透明域をもちます.
ムネエソは,ワニトカゲギス目ムネエソ科に属する全長5cm程の中深層性魚類です.本種は世界の熱帯から温帯水域にかけて広く分布し,水深
200 mから3,000 m に生息します.日本では北海道から沖縄にかけての太平洋岸沖に出現します.本科はキュウリエソ亜科とムネエソ亜科に分類され,日本周辺からは6属17種が報告されています.ムネエソ亜科は,眼が大きいこと,体が小判状で高く短いこと,体側下方に発光器が並ぶことで他の本目魚類との識別は容易です.本亜科の中でも,ムネエソ属
Sternoptyx は,他のテンガンムネエソ属 Argyropelecus やホウネンエソ属
Polyipnus に比べると,体全体が斜め後方へ傾いたように変形し(四角形から平行四辺形に),臀鰭基底部に透明域をもちます.
{kind=link}
ムネエソ科魚類は,他のワニトカゲギス目やハダカイワシ目,ヒメ目などの中深層性魚類と同様に,わずかに光が届く中深層で自身の影を消すために発光器を使います.これはカウンターシェーディングと呼ばれています.背面が黒く体側が銀色の体と発光器からの弱い光(生息水深に達する光と同量)により,隠れる場所のない中深層で捕食者の眼を欺くことができます.
参考文献
Aizawa, M. 2002. 96. Sternoptychidae, marine hatchetfishes.
Pages 311-317, 1475-1476 in T. Nakabo, ed. Fishes of Japan
with pictorial keys to the species English edition. Tokai University
Press, Tokyo.
写真個体:BSKU
51870, ca. 40 mm SL,2000年8月26日,土佐湾中央部,中央水産研究所調査船こたか丸,オッタートロール,水深800m.
(遠藤広光)
2004年 10月
ギマ Triacanthus
biaculeatus (Bloch,
1786)(フグ目ギマ科) ギマはフグ目ギマ科の魚で,インド・西太平洋の沿岸域に生息しています.ギマ科魚類は世界で7種が確認されていますが,日本に分布する種はギマのみです.ギマという魚は,あまりなじみがないかもしれませんが,浜名湖や三河湾沿岸では食用として漁獲されています.高知県では「つのはぎ」と呼ばれていますが,稀に釣りや底曳き網で漁獲される程度で食用としての価値はあまりありません.ギマは底曳き網で漁獲される時にはまとまって捕られることから,群れを形成していると考えられます.
ギマはフグ目ギマ科の魚で,インド・西太平洋の沿岸域に生息しています.ギマ科魚類は世界で7種が確認されていますが,日本に分布する種はギマのみです.ギマという魚は,あまりなじみがないかもしれませんが,浜名湖や三河湾沿岸では食用として漁獲されています.高知県では「つのはぎ」と呼ばれていますが,稀に釣りや底曳き網で漁獲される程度で食用としての価値はあまりありません.ギマは底曳き網で漁獲される時にはまとまって捕られることから,群れを形成していると考えられます.
従来,ギマ科はベニカワムキ科と共に原始的なフグ目魚類とされていました.ところが,最近の研究では,ベニカワムキ科よりもモンガラカワハギ科やカワハギ科に近い仲間ではないかという説が唱えられました.ギマ科は,腹鰭に棘があること,顎が前方に突出可能であること等がベニカワムキ科と同様ですが,頭の骨や肩帯の骨がモンガラカワハギ科と同様の状態を示すようです.ギマ科の鱗はベニカワムキ科と似ています.しかしながら生息場所はベニカワムキ科よりもモンガラカワハギ科,カワハギ科と似ています.さて,味はどちらに似ているのでしょうか?幸い,フグ科の魚と違って,ギマ科やベニカワムキ科,モンガラカワハギ科(一部シガテラ毒に注意!),カワハギ科の魚にはフグ毒はないとされています.どなたか食べ比べをしてみませんか?新たな発見があるかも知れません.
{kind=link}
参考文献
加納碩雄.1988.脊椎動物におけるフグ毒の分布.橋本周久(編),pp.
32-44,フグ毒研究の最近の進歩.恒星社厚生閣,東京.
Santini F,
Tyler JC. 2002. Phylogeny and
biogeography of the extant species of triplespine fishes (Triacanthidae,
Tetraodontiformes). Zoologica Scripta 31: 321-330
Santini F,
Tyler JC. 2003. A phylogeny
of families of fossil and extant tetraodontiform fishes (Acanthomorpha,
Tetraodontiformes), Upper Cretaceous to Recent. Zool J Linn Soc
139:565-617
鈴木克美・日置勝三・北沢博.1983.ギマ Triacanthus biaculeatus (フグ目ギマ科)の水槽内産卵と生活史.東海大学紀要海洋学部
17: 131-138
Tyler JC. 1980. Osteology, phylogeny, and higher
classification of the fishes of the order Plectognathi (Tetraodontiformes).
NOAA Tech Rep NMFS Circular 434:1-42
Winterbottom
R. 1974. The familial phylogeny
of the Tetraodontiformes (Acanthopterygii: Pisces) as evidenced
by their comparative myology. Smithson Contrib Zool 155:1-201
写真個体:BSKU 61019, ca.20 cm SL, 2002年11月15日,高知県幡多郡佐賀町佐賀漁港
他の写真個体:BSKU
58995, 2002年6月12日,高知県幡多郡佐賀町佐賀漁港
{kind=link}
(中江雅典)
2004年 9月
リュウグウベラギンポ Trichonotus
elegans Shimada
et Yoshino, 1984(スズキ目ベラギンポ科) ベラギンポ科魚類はベラギンポ属のみを含み,インド-太平洋海域から9種が知られ,そのうち日本にはベラギンポ,クロエリギンポ,リュウグウベラギンポの3種が分布しています.ベラギンポ科魚類は、水深5-50mの砂地に生息し,ハーレムを作ることが知られ、雄は最大で全長20
cm まで成長し、雌は雄よりも小さく,体色や鰭の長さなど明らかな雌雄差が見られます。
ベラギンポ科魚類はベラギンポ属のみを含み,インド-太平洋海域から9種が知られ,そのうち日本にはベラギンポ,クロエリギンポ,リュウグウベラギンポの3種が分布しています.ベラギンポ科魚類は、水深5-50mの砂地に生息し,ハーレムを作ることが知られ、雄は最大で全長20
cm まで成長し、雌は雄よりも小さく,体色や鰭の長さなど明らかな雌雄差が見られます。
リュウグウベラギンポは,沖縄県八重山諸島の鳩間島から採集された標本をもとに記載されました.琉球諸島の水深10-30mの砂地での生息が確認されていますが,インドネシアやインド洋のモルジブ諸島の沿岸でも水中写真が撮影されています。高知県沖の島では今回初めて標本を採集したので,南日本での分布の北限記録となります。本種は紅海に分布するT.nikiiによく似ますが,体側線前方の上下に無鱗域があることで,本科の他種全てと区別できます。また、本種の雄では背鰭棘の3本が長く伸長すること、雌では背鰭前端に白く縁取られた黒斑をもつことが特徴です(雌雄の写真はこちら)。日本産の他種に比べ,水中では体色が薄く,全体的に白く見えます.海底直上では潮の流れに対して,その場でホバリングするように遊泳します。リュウグウベラギンポの和名は,その美しい体色と細長い体を波打たせて遊泳する行動に由来します。
リュウグウベラギンポのハーレムは,1個体の雄に対し,約12個体の雌によって構成されます。また,ハーレムに違う雄が入ってくると,ディスプレイによる威嚇が行われます。さらに,本種はメスからオスへと性転換をすることが分かっています。
英語名でサンドダイバーと呼ばれるベラギンポ科魚類は,敵が近づくと頭からすばやく砂に潜り,頭部先端だけを出し様子をうかがう行動を見せます。その行動は種によって違いがあり、リュウグウベラギンポは他種に比べあまり砂に潜らず,泳いで逃げることの方が多いようです。
{kind=link}
引用文献
Shimada,K.and
Yoshino,T. 1984 A new trichonotid
fish from the Yaeyama Islands, Okinawa prefecture, Japan. Jpn.J.Ichthyol.15-19.
吉野哲夫・島田和彦. 1984. ベラギンポ科. 益田一・尼岡邦夫・荒賀忠一・上野輝彌・吉野哲夫(編).pp. 279-280,
日本産魚類大図鑑.東海大学出版会,東京.
岡村 収.1997.ベラギンポ科.Trichonotidae.
岡村 収・尼岡邦夫(編),pp. 328,日本の海水魚,山と渓谷社,東京.
写真個体:BSKU 71966, 129.4 mm SL, オス,2004年7月19日,高知県宿毛市沖の島,水深12m.雌雄の写真はこちら(メス 91.6 mm
SL).
(片山英里)
2004年 8月
アキアナゴ Gorgasia taiwanensis
Shao, 1986(ウナギ目アナゴ科)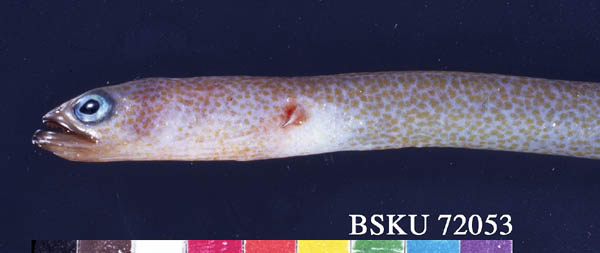 英語名ではガーデンイールと呼ばれるアナゴ科チンアナゴ亜科魚類には,世界中の熱帯から亜熱帯の浅海域に生息するシンジュアナゴ属
Gorgasia とチンアナゴ属 Heteroconger の2属26種が知られています(Castle
and Randall, 1999).そのうち日本には,前属のシンジュアナゴ G. japonicus,ニシキアナゴ
G. preclara,アキアナゴ G. taiwanensis,後属のチンアナゴ H.
hassi およびゼブラアナゴ H. polyzona の計5種が分布します (Nakabo, 2002).ガーデンイールは,浅海域(多くはサンゴ礁域)の砂地に巣穴を掘り,その穴から半身を出して,潮に流されて来る餌生物を捕食します.通常はコロニーを形成するため,多くの個体が穴から身を乗り出した状態は,まるで「アスパラ畑」のようです.この仲間は極めて臆病なため,ダイバーが近づくとすぐに穴の奥深くに入ってしまいます.
英語名ではガーデンイールと呼ばれるアナゴ科チンアナゴ亜科魚類には,世界中の熱帯から亜熱帯の浅海域に生息するシンジュアナゴ属
Gorgasia とチンアナゴ属 Heteroconger の2属26種が知られています(Castle
and Randall, 1999).そのうち日本には,前属のシンジュアナゴ G. japonicus,ニシキアナゴ
G. preclara,アキアナゴ G. taiwanensis,後属のチンアナゴ H.
hassi およびゼブラアナゴ H. polyzona の計5種が分布します (Nakabo, 2002).ガーデンイールは,浅海域(多くはサンゴ礁域)の砂地に巣穴を掘り,その穴から半身を出して,潮に流されて来る餌生物を捕食します.通常はコロニーを形成するため,多くの個体が穴から身を乗り出した状態は,まるで「アスパラ畑」のようです.この仲間は極めて臆病なため,ダイバーが近づくとすぐに穴の奥深くに入ってしまいます.
高知県では大月町柏島からアキアナゴとチンアナゴの2種が報告され,沖の島ではアキアナゴの生態写真が撮影されました(平田ほか,1996;浅野,1997).アキアナゴは,台湾で採集された標本に基づき1986年に新種として記載され,日本では西表島,高知県(柏島と沖の島),駿河湾から記録されています.
引用文献
浅野博利.1997.アナゴ科.Congridae. 岡村 収・尼岡邦夫(編),pp. 84-88,日本の海水魚,山と渓谷社,東京.
Castle,
P.H.J. and J.E. Randall. 1999. Revision of Indo-Pacific garden eels (Congridae:
Heterocongrinae), with descriptions of five species. Indo-Pacific
fishes, (30): 1-52.
平田智法・山川 武・岩田明久・真鍋三郎・平松 亘・大西信弘.1996.高知県柏島の魚類相.行動と生態に関する記述を中心として.Bull. Mar. Sci. Fish.,
Kochi Univ., 16: 1-177.
Nakabo, T.
2002. Fishes of Japan with pictorial keys to the species English
edition. Tokai University Press, Tokyo. 1xi+1749 pp.
写真個体:BSKU 72053, 479 mm TL, 2004年7月21日,高知県宿毛市沖の島,水深10m (採集者:井本善次),全体写真はこちら.
チンアナゴ Heteroconger
hassi (Klausewitz et Eible-Eibesfeldt, 1959): BSKU 72054,
313 mm TL, 高知県宿毛市沖の島,水深10m(採集者:遠藤広光),頭部写真,全体写真はこちら.
{kind=link}
{kind=link}
{kind=link}
(遠藤広光)
2004年 7月
ナガハダカ Symbolophorus
californiensis (Eigenmann et Eigenmann,
1889)(ハダカイワシ目ハダカイワシ科) ハダカイワシ科魚類 (Myctophidae) は,北極海を除く世界の大洋に広く分布し,水深200~1200mの中深層に生息しています
(Nelson,1994).また,本科魚類はマイクロネクトンと呼ばれる小型遊泳動物の中で,ヨコエソ科と並び生物量の多い動物群です.現在,本科魚類は世界で30属約250種が知られ,日本近海からは23属88種が報告されています(中坊,2002).本科魚類には,夜間に表層近くまで浮上する日周鉛直移動型と,浮上せず中深層に生息する非移動型が知られています.また,夜間表層に浮上する本科魚類の生物量は数億トン程度と見積もられ,イカ・タコ類,大型魚類,海鳥や海獣などの重要な餌となっています
(川口,1998).一方,昼間には水深200~1000mの中深層で,多くの底魚や浮魚に捕食されており,本科魚類は表層の有機物を中深層まで運ぶ,重要な橋渡し役を担っているといえます.
ハダカイワシ科魚類 (Myctophidae) は,北極海を除く世界の大洋に広く分布し,水深200~1200mの中深層に生息しています
(Nelson,1994).また,本科魚類はマイクロネクトンと呼ばれる小型遊泳動物の中で,ヨコエソ科と並び生物量の多い動物群です.現在,本科魚類は世界で30属約250種が知られ,日本近海からは23属88種が報告されています(中坊,2002).本科魚類には,夜間に表層近くまで浮上する日周鉛直移動型と,浮上せず中深層に生息する非移動型が知られています.また,夜間表層に浮上する本科魚類の生物量は数億トン程度と見積もられ,イカ・タコ類,大型魚類,海鳥や海獣などの重要な餌となっています
(川口,1998).一方,昼間には水深200~1000mの中深層で,多くの底魚や浮魚に捕食されており,本科魚類は表層の有機物を中深層まで運ぶ,重要な橋渡し役を担っているといえます.
ナガハダカは,日本の太平洋岸沖では北海道から駿河湾,土佐湾にかけて出現し,おもに北太平洋の亜寒帯水と中央水の遷移帯までの冷水域に分布します(川口,1998).ナガハダカ属
(Symbolophorus) のうち,本種はマガリハダカ (S. evermanni) に最も類似しますが,臀鰭基底後部の上方に4~5個
(稀に3個) の後部臀鰭発光器(AOp)があることで,区別されています.また,体側後部発光器(Pol)は脂鰭起部よりかなり前にあることや,第一肛門上発光器(SAO1)は第二腹部発光器(VO2)の直上,あるいはそれよりわずかに後方にあること,前部臀鰭発光器(AOa)は一直線上にあること,口蓋骨歯は絨毛状で狭歯部をなし,肥大歯がないなどの特徴をもちます.本種は本属の中では大型種で,体長約11cmにまで成長します.日周鉛直移動型の本種は,夜間に小型甲殻類プランクトンを捕食するため水深
50 m 以浅まで浮上します.本標本は,高知県室戸市にある椎名漁港(室戸岬東側)の定置網で漁獲されました.水深はおよそ100mに設置されているこの定置網では,室戸岬東部沿岸の急深の海底地形と海流のため,夜間に浮上する中深層性魚類も沿岸性魚類に混じり漁獲されます.
引用文献
Eigenmann, C.H. and
R.S. Eigenmann. 1889. Notes from the San
Diego Biological Laboratory. The fishes of Cortez Banks.West Amer.Sci.6 (48): 123-132.
川口弘一.1998.ハダカイワシ目.中坊徹次・望月賢二(編),pp.55-57.日本動物大百科.平凡社,東京.
益田一・尼岡邦夫・荒賀忠一・上野輝彌・吉野哲夫(編).1984.日本産魚類大図鑑.東海大学出版会,東京.xx+448 pp., 370 pls.
Nakabo, T. 2002. Fishes of Japan with pictorial
keys to the species English edition. Tokai University Press,
Tokyo. 1xi+1749 pp.
Nelson, J.S. 1994. Fishes
of World. Third edition. John Wiley & Sons, Inc., New York.
xvii+600 pp.
写真標本データ:BSKU 70422,105.3
mm SL,2004年4月29日,高知県室戸市椎名漁港(採集&写真:三宅崇智).白バックはこちら
{kind=link}
(三宅崇智)
2004年 6月
ソコヌメリ Bathycallionymus
sokonumeri (Kamohara,
1936)(スズキ目ネズッポ科) ソコヌメリは南日本太平洋岸沖の水深150~200mの大陸棚縁辺域に生息するネズッポ科魚類です.日本産のトンガリヌメリ属
Bathycallionymus は,本種とトンガリヌメリ B. kaianus Gunther
, クジャクソコヌメリB. formosanus (Fricke) の3種のみです.本種は第1背鰭の第1棘が短いこと(第2棘とほぼ同長),雄では第2背鰭前上方先端に黒色斑がないことで他種と区別されています.
ソコヌメリは南日本太平洋岸沖の水深150~200mの大陸棚縁辺域に生息するネズッポ科魚類です.日本産のトンガリヌメリ属
Bathycallionymus は,本種とトンガリヌメリ B. kaianus Gunther
, クジャクソコヌメリB. formosanus (Fricke) の3種のみです.本種は第1背鰭の第1棘が短いこと(第2棘とほぼ同長),雄では第2背鰭前上方先端に黒色斑がないことで他種と区別されています.
ソコヌメリは1936年に蒲原稔治博士により Callionymus
sokonumeri として新種記載されました.この記載に使われた全長173 mm のホロタイプ標本は,土佐湾の水深およそ150m(80尋)で漁獲され,高知市御畳瀬魚市場に水揚げされたものでした.ところが,このタイプ標本は第二次世界大戦時の空襲(1945年)により焼失してしまいます.蒲原博士は,戦後ふたたび採集した標本をもとに失われたタイプ標本の種についてネオタイプ指定を行いました(現在の動物の学名命名規約と照らし合わせると無効).しかし,この時点で土佐湾から新たなソコヌメリの標本は採集されていませんでした.その後,ソコヌメリはNakabo
(1982) が設立した新属のBathycallionymus に含められました.一方,Fricke (1983)
はこの属を独立属とは認めず,ネズッポ属Callionymus の亜属としています(Bathycallionymus
はFricke の C. kaianus 種群に相当).また,近年日本ではネズッポ属に Repomucenus
が使われています.これは現在Callionymus に属する種が地中海からヨーロッパの沿岸に分布しており,過去にCallionymus
にひとまとめにされていた太平洋やインド洋産の種が別属に移されたためです.ネズッポ科魚類の分類体系,亜属や属の定義について,中坊徹次博士とRonald
Fricke博士(ドイツの魚類学者)の一連の研究による見解の相違は極めて大きいことから,現在2通りの分類体系が使用され混乱を招いています.日本では,多くの骨格系の形態形質を用いた中坊博士の系統仮説に基づく分類体系に従っています.
参考文献
Fricke, R. 1983.
Revision of the Indo-Pacific genera and species of the dragonet
family Callionymidae (Teleostei). Theses Zool. v. 3: 1-774.
Kamohara,
T. 1936. Two new deepsea fishes
from Japan. Annot. Zool. Jpn. v. 15 (no. 4): 446-448.
Nakabo, T. 1982. Revision of genera of the dragonets (Pisces: Callionymidae).
Publ. Seto Mar. Biol. Lab. v. 27 (no. 1/3): 77-131.
写真標本データ:2004年3月8日,高知市御畳瀬魚市場(大手繰),122 mm SL,白バックの写真はこちら
{kind=link}
(遠藤広光)
2004年 5月
オニソコホウボウ Pterygotrigla
multiocellata (Matsubara,
1937)(スズキ目フサカサゴ亜目ホウボウ科) オニソコホウボウは,南日本の太平洋岸沖,東シナ海,南シナ海,九州-パラオ海嶺,ニューカレドニアに分布し,水深220~350mの海底に生息します.本種は吻突起が長く強大で,その起部側面と鼻骨にそれぞれ1対の小棘をもつこと,第2背鰭基底に骨質板がないなどの特徴をもちます.日本産のホウボウ科魚類の中では,ホウボウ
Chelidonichthys spinosus に次ぐ大型種で,全長35
cm 程度になります.
オニソコホウボウは,南日本の太平洋岸沖,東シナ海,南シナ海,九州-パラオ海嶺,ニューカレドニアに分布し,水深220~350mの海底に生息します.本種は吻突起が長く強大で,その起部側面と鼻骨にそれぞれ1対の小棘をもつこと,第2背鰭基底に骨質板がないなどの特徴をもちます.日本産のホウボウ科魚類の中では,ホウボウ
Chelidonichthys spinosus に次ぐ大型種で,全長35
cm 程度になります.
Matsubara (1937) は,1937年に熊野灘の水深275mで採集された1個体の標本に基づき,新属新種 Parapterygotrigla multiocellata オニソコホウボウを記載しました.その後,このオニソコホウボウ属 Parapterygotrigla は ソコホウボウ属 Pterygotrigla Waite, 1899 のシノニムとされたり,独立の属とされたり,研究者により扱いが異なります(例えば,Del Cerro and Lloris, 1997; Richards, 1999).本種が記載された翌年に,Kamohara (1938) はオニソコホウボウをソコホウボウ属に含めています.おそらくこの分類学的処置には納得しなかったであろう松原博士は,その後「魚類の形態と検索(松原,1955)」の中で,オニソコホウボウ属を有効とし,逆にKamohara (1936) が記載したハナナガソコホウボウ Pterygotrigla macrorhynchus を,オニソコホウボウ属へ含めています.このグループに深く係わった両魚類学者の見解の違いが,良く現れている点と言えるでしょう.最近の文献を見ると,両種共にソコホウボウ属へ含める研究者が多いようです.
参考文献
Del Cerro, L. and D. Lloris. 1997. Gurnard fishes (Scorpaeniformes,
Triglidae) from off New Caledonia, with description of five new
species. In: Seret, B. (ed.), Resultats des Campagnes MUSORSTOM,
Volume 17. Mem. Mus. natn. Hist. nat., 174: 91-124.
Kamohara, T. 1936. On a new fish of the Triglidae from
Kochi, Japan. Dobutsugaku Zasshi [Zool. Mag. Tokyo] v. 48 (nos.
8-10): 481-483. [In Japanese, English summ.]
Kamohara, T. 1938. On the offshore bottom-fishes of
Prov. Tosa, Shikoku, Japan. pp. 1-86. Fishes Prov. Tosa. Maruzen
Co., Tokyo.
Matsubara, K. 1937.
Studies on the deep-sea fishes of Japan. V. Diagnosis of a new
mail-cheeked fish, Parapterygotrigla multiocellata n.
g., n. sp., belonging to Triglidae. Zool.Mag. (Japan), 49(7):
266-267.
松原喜代松.1955.魚類の形態と検索 II.石崎書店,東京,
vi+791-1605 pp.
Richards, W.J.
1999. Triglidae. Pages 2359-2382 in K.E. Carpenter and
V.H. Niem, eds. FAO species identification guide for fishery
purposes. The living marine resources of the Western Central
Pacific. Volum 4. Bony fishes part 2 (Mugilidae to Carangidae).
FAO, Rome.
写真標本データ:BSKU
68696, 255 mm SL, 2004年2月19日,高知市御畳瀬魚市場(採集者:黒木広大・平松 亘)
(遠藤広光)
2004年 4月
ムツエラエイ Hexatrygon
bickelli Heemstra
et Smith(エイ目ムツエラエイ科)他の写真はこちら
 ムツエラエイは,エイ目の中では採集例が極めて少ない深海性の種です.本種はインド-西部太平洋海域に広く分布し,大陸棚上部の水深
120-1120 mで採集されています.今回の標本は土佐湾西部沿岸の小型底曳き網漁で漁獲され,その水深はおよそ120mでした.しかし,1997年5月に伊豆大島の水深38mで遊泳する雌個体がNHKの潜水取材班により撮影され(石原ほか,1998),また駿河湾沿岸の大瀬崎でも,2000年6月に水深15~20mで遊泳時の写真が撮影されています(瀬能・原,2000).そのため,本種は浅海域と深海域を移動するという可能性も考えられます.日本では,沖縄舟状海盆の水深
710 m(Ishihara and Kishida, 1984),東シナ海大陸斜面の水深 500 m(山田,1988),小田原市沖の相模湾の水深120m(瀬能・原,2000),和歌山県沿岸の水深38m(未発表)で各1個体が採集されています.日本での採集記録として,正式に公表された標本は3個体のみです.しかし,世界では未報告を含めると,合計20個体以上のムツエラエイ類が採集または観察されています(石原,1996).
ムツエラエイは,エイ目の中では採集例が極めて少ない深海性の種です.本種はインド-西部太平洋海域に広く分布し,大陸棚上部の水深
120-1120 mで採集されています.今回の標本は土佐湾西部沿岸の小型底曳き網漁で漁獲され,その水深はおよそ120mでした.しかし,1997年5月に伊豆大島の水深38mで遊泳する雌個体がNHKの潜水取材班により撮影され(石原ほか,1998),また駿河湾沿岸の大瀬崎でも,2000年6月に水深15~20mで遊泳時の写真が撮影されています(瀬能・原,2000).そのため,本種は浅海域と深海域を移動するという可能性も考えられます.日本では,沖縄舟状海盆の水深
710 m(Ishihara and Kishida, 1984),東シナ海大陸斜面の水深 500 m(山田,1988),小田原市沖の相模湾の水深120m(瀬能・原,2000),和歌山県沿岸の水深38m(未発表)で各1個体が採集されています.日本での採集記録として,正式に公表された標本は3個体のみです.しかし,世界では未報告を含めると,合計20個体以上のムツエラエイ類が採集または観察されています(石原,1996).
ムツエラエイは,和名や英名(sixgill stingray) の由来となった体盤腹面にある6対の鰓孔(他のエイ目魚類ではすべて5対,ただしムツエラエイも鰓弓は5対)と尾部の棘,退化的で極めて小さい脳,長く大きな吻などにより特徴づけられます(写真個体では,棘は根元から折れ,尾鰭は欠如している).また,体は全体的に寒天質の部分が多く,本種を「エイの中のシャチブリ」と呼んでもよい程です.長い吻にはかなりの柔軟性があり,下面には餌の探査に使われる電気受容器のロレンチニ瓶がよく発達します.頭部下面にある口は,下方へ著しく突出可能なことから,吻の特徴とともに底生性動物の捕食に優れていると考えられます.今回採集した標本を調べたところ,胃の中は空でした.
{kind=link}
{kind=link}
{kind=link}
エイ目トビエイ亜目のムツエラエイ科には,現在ムツエラエイ属 Hexatrygon のみが含まれています.Heemstra and Smith (1980) は,南アフリカ南東部のポートエリザベスの海岸(インド洋に面する)に流れ着いた雌の1個体の標本(全長は105 cm)に基づき,その詳しい骨格の解剖学的特徴とともに H. bickelli を新亜目新科新属新種として発表しました.種小名は発見者の Dave Bickell 氏に献名されたものです.それ以降,台湾周辺から採集された標本により,H. longirostra (Chu & Meng, 1986),H. yangi Shen & Liu, 1984,H. brevirostra Shen, 1986,H. taiwanesis Shen, 1986 の4種が次々と記載されました.これらの研究では,おもに吻の長さや形状の違いが識別形質とされました.しかし,どの種も1~2個体の標本に基づくため,形態の種内変異は考慮されませんでした.その後 Compagno and Last (1999) は,成長に伴う吻の伸長,標本の固定や保存により吻が変形する可能性を指摘し,H. bickelli のみを有効種として,他の4種を本種のシノニムとみなしています.本稿では,この見解に従い,ムツエラエイの学名を H. bickelli としました.
McEachran et al. (1996) は,エイ類全体の形態学的情報を集め,エイ上目の分岐仮説と分類体系を示しました.その中で,ムツエラエイは,トビエイ目トビエイ亜目の中で,最初に分岐するグループとされています(アカエイ類+ツバクロエイ類+トビエイ類のクレード[単系統群]の姉妹群).また,近年行われたトビエイ類に関するいくつかの系統学的研究でも,ほぼ同様に初期派生群と推測されています(石原ほか,1999).これの仮説を裏付ける生態的特徴として,本種の遊泳方法が挙げられます.派生的なトビエイ類は,例えばオニイトマキエイ(マンタ)のように,体盤の両翼を羽ばたいて遊泳します.一方,ムツエラエイは,体盤側部を波打たせて遊泳するため,トビエイ類の中では原始的と考えられます(石原ほか,1998).
参考文献
Compagno, L. J. V. and P. R. Last. 1999. Hexatrygonidae. Pages 1477-1478
in K.E. Carpenter and V.H. Niem, eds. FAO species identification
guide for fishery purposes. The living marine resources of the
Western Central Pacific. Volum 3. Batoid fishes, Chimaeras and
bony fishes part 1 (Elopidae to Linophrynidae). FAO, Rome.
Heemstra,
P. C. and M. M. Smith. 1980.
Hexatrygonidae. A new family of stingrays (Myliobatiformes: Batoidea)
from South Africa, with comments on the classification of batoid
fishes. Ichthol. Bull. J.L.B. Smith Inst. Ichthyol., 43: 1-17.
石原 元.1996.ムツエラエイ類.千石正一・疋田努・松井正文・仲谷一宏(編).pp. 172-173,日本動物大百科5 両生類・爬虫類・軟骨魚類.平凡社,東京.
Ishihara,
H. and S. Kishida. 1984. First
records of the sizgill stingray Hexatrygon longirostra
from Japan. Japan. J. Ichthyol., 30 (4): 452-454.
石原 元・本間公也・南方盈進・木原英雄・池田信浩・秋吉一朗・川瀬直也.1998.伊豆大島浅海域におけるムツエラエイの出現.板鰓類研究会報,(34): 17-19.
石原 元・西田清徳・本間公也.1999.板鰓亜綱エイ目内及び各亜目内の系統類縁関係に関する最近の研究のレビュー.月刊海洋
号外,(16): 31-44.
Last, P. R. and J. D. Stevens. 1994. Sharks and rays of Australia. Australia,
CSIRO, 512 pp.
McEachran, J.D.K. Dunn
and T. Miyake. 1996. Interrelationships
of the batoid fishes (Chondrichthyes, batoidea). Pages 63-84
in M. L. J. Stiassny, L. R. Parenti, and G. D. Johnson
(eds), Interrelationships of fishes. Academic Press, San Diego.
瀬能 宏・原 真一.2000.今月の魚 ムツエラエイ Hexatrygon
longirostra (Chu et Meng, 1981).I. O. P. Diving News(伊豆海洋公園通信),11 (9): 1.
Smith, M. M. and P. C.
Heemstra. 1995. Family No.
31: Hexatrygonidae. Pages 142-143 in M. M. Smith and P.
C. Heemstra, eds. Smith's sea fishes. J. L. B. Smith Institute
of Ichthyology, Grahamstown. (初版第3刷)
山田梅芳.1988.ムツエラエイ属の1種 Hexatrygon
yangi Shen et Liu.西海区水産研究所ニュース,(59):
1.
写真標本データ:BSKU
87842, 103+ cm TL. 2004年4月1日,高知県幡多郡佐賀町佐賀漁港で採集(採集者:町田吉彦ほか,写真撮影:遠藤広光)
*土佐湾のムツエラエイの記録が論文になりました(2005/06/10)
Endo, H.
and Y. Machida. 2005.
Sixgill stingray Hexatrygon bickelli collected from Tosa
Bay (Rajiformes: Hexatrygonidae).
Bull. Shikoku
Inst. Nat. Hist., 2: 51-57.(四国自然史科学研究)
(遠藤広光)
2004年 3月
チカメエチオピア Eumegistus
illustris Jordan
et Jordan (スズキ目シマガツオ科 )
 シマガツオ科には,世界の熱帯から温帯海域に分布する7属約20種が知られています.そのうち,チカメエチオピア属 (Eumegistus) は,太平洋産のチカメエチオピアと大西洋産の
E. brevorti の2種のみを含みます.チカメエチオピアは,本科の中では稀種で,ハワイ諸島(基産地)やポリネシア,南日本沿岸の相模湾(真鶴),土佐湾(御畳瀬魚市場),九州-パラオ海嶺,沖縄島での採集記録があります.本科の多くの種は,外洋の水深
200 m 以浅の表層域に群で生息します.しかし,チカメエチオピアは水深 275~620
mで記録されており,本科の中では最も深い水深帯に出現します.また,シマガツオ Brama japonica
Hilgendorf などは夜間には表層域へ浮上したり,南北への季節的な回遊を行うことが知られています.
シマガツオ科には,世界の熱帯から温帯海域に分布する7属約20種が知られています.そのうち,チカメエチオピア属 (Eumegistus) は,太平洋産のチカメエチオピアと大西洋産の
E. brevorti の2種のみを含みます.チカメエチオピアは,本科の中では稀種で,ハワイ諸島(基産地)やポリネシア,南日本沿岸の相模湾(真鶴),土佐湾(御畳瀬魚市場),九州-パラオ海嶺,沖縄島での採集記録があります.本科の多くの種は,外洋の水深
200 m 以浅の表層域に群で生息します.しかし,チカメエチオピアは水深 275~620
mで記録されており,本科の中では最も深い水深帯に出現します.また,シマガツオ Brama japonica
Hilgendorf などは夜間には表層域へ浮上したり,南北への季節的な回遊を行うことが知られています.
チカメエチオピアは,アメリカ合衆国の高名な魚類学者であるDavid Starr Jordan が,1921年8月にハワイのオアフ島のホノルルに滞在した際に市場で購入した1個体の標本を基に,新属新種として記載されました(Jordan and Jordan, 1922).その原記載には,ホロタイプの体長が約 60 cm(2フィート),重さは4キロ弱(9ポンド近く)で,購入時に半身の肉はすでになかったとの記述がありました(ホロタイプ原図の白色部分).シマガツオ科魚類は,成長により体のプロポーション,鰭の形や長さなどがかなり変化することが知られています.本種のホロタイプの原図を見ると,体型や尾鰭の形は写真個体と随分異なり,ヒレジロマンザイウオの尾鰭のようです.ホロタイプが成熟個体だとすると,本種の尾鰭は成長に伴い後縁中央の張り出し部分が次第に短くなり,ほとんど目立たなくなって,その縁辺も白くなることが考えられます.それとも, Jordan and Jordan(1922)が記載した種と私たちがチカメエチオピアとしている種が異なっているのでしょうか?
{kind=link}
{kind=link}
チカメエチオピアの「エチオピア」は,シマガツオに対して用いられた地方名です.その名前は,昭和9年(1934年)にエチオピアの皇太子が日本へお嫁さんを捜しに来た頃,相模湾でシマガツオが大量に漁獲されたことに由来するようです.その後,三浦半島周辺や東京の市場などで広く使われるようになり,チカメエチオピアやツルギエチオピアの標準和名の中に登場しています.
Fishbase に掲載されているチカメエチオピアの写真(J. E. Randall 撮影: 50 cm
SL, 63 cm TL)はこちら
参考文献
Jordan, D. S. and
E. K. Jordan. 1922. A list
of the fishes of Hawaii, with notes and descriptions of new species.
Mem. Carnegie Mus., 10 (1): 1-92, 4 pls.
Last, P. R. and M. Moteki. 2001. Bramidae. Pages 2824-2836 in
K.E. Carpenter and V.H. Niem, eds. FAO species identification
guide for fishery purposes. The living marine resources of the
Western Central Pacific. Volum 5. Bony fishes part 3 (Menidae
to Pomacentridae). FAO, Rome.
望月賢二.1984.シマガツオ科.益田一・尼岡邦夫・荒賀忠一・上野輝彌・吉野哲夫(編).pp. 154-155,日本産魚類大図鑑.東海大学出版会,東京.
谷津明彦.1997.シマガツオ科.Bramidae. 岡村 収・尼岡邦夫(編),pp. 328,日本の海水魚,山と渓谷社,東京.
写真標本データ:BSKU 68721, 40 cm SL. 2004年2月20日,高知県高知市御畳瀬魚市場で採集(採集:野川悠一郎・遠藤広光;撮影:野川悠一郎).青バックの標本写真はこちら
{kind=link}
(遠藤広光)
2004年 2月
キュウシュウヒゲ Caelorinchus
jordani Smith et
Pope (タラ目ソコダラ科 )
 ソコダラ科は,深海域で多様に種分化したタラ目最大のグループで,現在およそ35属約350種あまりを含んでいます.本科魚類の多くは底生性で,大陸棚の水深150m前後から大陸斜面,水深6,500mの大洋の海底上に生息し,一部の種は中深層にも出現します.また,本科魚類は北極海の一部を除き世界大洋に広く分布します.
ソコダラ科は,深海域で多様に種分化したタラ目最大のグループで,現在およそ35属約350種あまりを含んでいます.本科魚類の多くは底生性で,大陸棚の水深150m前後から大陸斜面,水深6,500mの大洋の海底上に生息し,一部の種は中深層にも出現します.また,本科魚類は北極海の一部を除き世界大洋に広く分布します.
キュウシュウヒゲは,トウジン属 (genus Caelorinchus)の中では小型種で,全長25cm程度になります.本種は,南日本の太平洋沿岸から東シナ海にかけて分布し,水深およそ150mから400mの大陸棚縁辺から斜面上部に生息します.ソコダラ科の中では,本種は最も浅い水深に出現します.トウジン属は,どの種も頭部が大きく,吻がよく発達するのですが,本種は頭部が小さく,吻も短いことが特徴です.腹部には,胸鰭の前方から肛門まで黒色の発光器が伸びています.
Smith and Pope (1906)は,
第一著者の Hugh M. Smith が1903年6月16日に鹿児島で採集した1個体の標本に基づき,キュウシュウヒゲを新種として記載しました.和名は,その後に基産地の鹿児島に因んで付けられたようです.
参考文献
Jordan, D. S.,
S. Tanaka and J. O. Snyder.
1913. A catalogue of the fishes of Japan. J. Coll. Sci. Imp.
Univ. Tokyo, 33(1): 1-497, 396 figs.
Nakabo, T. 2002. Macrouridae. Pages 417-435,
1491 in T. Nakabo, ed. Fishes of Japan with pictorial
keys to the species, English edition.Tokai University, Press,
Tokyo.
Okamura,
O. 1970. Fauna Japonica, Macrourina
(Pisces). Academic Press of Japan, Tokyo. 216 pp, XLIV pls.
Smith,
H. M. and T. E. B. Pope. 1906.
List of fishes collected in Japan in 1913, with descriptions
of new genera and species. Proc. U.S. Nat. Mus., 31 (1489): 459-499,
12 figs.
写真標本データ:BSKU 87825, ca.20 cm TL. 2004年1月14日,高知県高知市御畳瀬魚市場で採集(撮影:藤原 暁)
(遠藤広光)
2004年 1月
トンガリセミホウボウ Dactyloptena
tiltoni Eschmeyer
(スズキ目セミホウボウ科 )
 本種は,セミホウボウ科の小型種で,フィリピンおよびオーストラリア北西岸沖の水深119~565mで採集された標本を基に,新種として記載されました(Eschmeyer,
1997).日本からは,土佐湾で採集された標本に基づき,先月初めて報告され,トンガリセミホウボウの和名が付けられています
(Sato et al., 2003).本種は,中央水産研究所こたか丸の底曵き網調査では土佐湾中央部のおよそ水深100~125m間で,また高知市御畳瀬魚市場の大手繰り網(およそ水深100~300m)や県西部の佐賀町(おそらく水深100m前後)で稀に採集されます.現在,セミホウボウ属
Dactyloptena には6種が知られ,本種は吻がよく尖ることで他種との識別は容易です.和名もその特徴に由来します(標本の背面写真はこちら).
本種は,セミホウボウ科の小型種で,フィリピンおよびオーストラリア北西岸沖の水深119~565mで採集された標本を基に,新種として記載されました(Eschmeyer,
1997).日本からは,土佐湾で採集された標本に基づき,先月初めて報告され,トンガリセミホウボウの和名が付けられています
(Sato et al., 2003).本種は,中央水産研究所こたか丸の底曵き網調査では土佐湾中央部のおよそ水深100~125m間で,また高知市御畳瀬魚市場の大手繰り網(およそ水深100~300m)や県西部の佐賀町(おそらく水深100m前後)で稀に採集されます.現在,セミホウボウ属
Dactyloptena には6種が知られ,本種は吻がよく尖ることで他種との識別は容易です.和名もその特徴に由来します(標本の背面写真はこちら).
セミホウボウ科は,その特異な形態学的特徴から,これまで独立の目(セミホウボウ目)とされることもありましたが,近年ではカサゴ目に含められてきました(例えば,Nakabo,
2002 ではカサゴ目セミホウボウ亜目).最近 Imamura (2000) は,本科の骨格や筋肉の状態を詳細に比較解剖して系統解析を行った結果,他のカサゴ目魚類との近縁性は低く,セミホウボウ科がスズキ目内においてアマダイ科およびキツネアマダイ科と単系統群をなすことを示しました.また,その分岐仮説では,本科はキツネアマダイ科の中のサンゴアマダイ属
(Hoplolatilus) と姉妹群関係にあり,これまでのキツネアマダイ科(3属を含む)は,側系統群であることが示されています.Imamura
(2000) は,この単系統群をセミホウボウ科としています.最近の真骨魚類の分子系統学的解析でも,セミホウボウ科は他のカサゴ目と離れた分岐に位置しています(Miya
et al., 2003).また,カサゴ目自体の多系統性も形態学および分子学的特徴の両面から示されました(Imamura
and Yabe, 2002; Miya et al., 2003).形態学的特徴からは,フサカサゴ亜目はハタ科と,カジカ亜目はゲンゲ亜目との近縁性がそれぞれ示唆されました(Imamura
and Yabe, 2002).カサゴ目はもはや実体のないグループとなってしまったようです.
{kind=link}
参考文献
Eschmeyer, W. N.
1997. A new species of Dactylopteridae
(Pisces) from the Philippines and Australia, with a brief synopsis
of the family. Bull. Mar. Sci., 60: 727-738.
Imamura, H. 2000. An alternative hypothesis on the phylogenetic
position of the family Dactylopteridae (Pisces: Teleostei), with
a proposed new classification. Ichthyol. Res., 47: 203-222.
Imamura,
H. and M. Yabe. 2002. Demise
of the Scorpaeniformes (Actinopterygii: Percomorpha): an alternative
phylogenetic hypothesis. Bull. Fish. Sci. Hokkaido Univ., 53
(3): 107-128.
Nakabo, T. 2002. Dactylopteridae. Pages 678, 1530 in T.
Nakabo, ed. Fishes of Japan with pictorial keys to the species,
English edition.Tokai University, Press, Tokyo.
Miya, M., H. Takeshima,
H. Endo, N.B. Ishiguro, J.G. Inoue, T. Mukai, T.P. Satoh, M.
Yamaguchi, A. Kawaguchi, K. Mabuchi, S.M. Shirai, and M. Nishida.
2003. Major
patterns of higher teleostean phylogenies: a new perspective
based on 100 complete mitochondrial DNA sequences. Mol. Phylogenet.
Evol., 26 (1): 121-138.
Sato, T., H. Endo, T. Nakabo and Y. Machida. 2003. First record of the helmet gurnard, Dactyloptena
tiltoni (Osteichthys: Dactylopteridae), from Japan with comments
on newly-recognized characters. Species Diversity, 8: 391-397.
写真標本データ:BSKU 66626,
ca. 11 cm SL. 2003年10月23日,高知県幡多郡佐賀町佐賀漁港.
(遠藤広光)