2011年12月の魚

エンドウヒメアンコウ Lophiodes endoi Ho et Shao, 2008 (アンコウ目アンコウ科)
エンドウヒメアンコウは1984年に出版された「沖縄舟状海盆及び周辺海域の魚類 I」の中で,ヒメアンコウ属の一種(Lophiodes sp.)として報告され,その後学名が付けられていませんでした(岡村,1984).本種は2008年に台湾のHusuan-Ching Ho 博士とKwang-Tsao Shao 博士により,土佐湾,沖縄舟状海盆,そして九州-パラオ海嶺,台湾,オーストラリアの南東岸と北西岸沖の水深261–750 mで採集された41標本に基づき記載されました.その後,ニューカレドニアからも報告されています(Ho et al., 2011).
Ho博士は,これまでに本研究室を3度訪問し,アンコウ目の標本を数多く調査しています.そのため,BSKUの8標本を本種のパラタイプに指定し,種小名には私の名前を付けてくれました.この論文は日本魚類学会の英文誌 Ichthyological Researchに掲載され,担当した編集委員が学名に従って,標準和名を付けています.著者が外国人で日本産の種を記載する場合には,担当編集委員が標準和名を考えて著者に提案し,学名とともに付けることがあります.
アンコウ目アンコウ科は4属24種を含み,そのうちヒメアンコウ属は15種を含む最大のグループです(Ho and Shao, 2008; Ho et al., 2009, 2011; Eschmeyer and Fong, 2011).本属の体サイズは,食用となる他属のアンコウ Lophiomus setigerus (約70 cm)やキアンコウ Lophius litulon (約1 m)に比べると小さく,本種の最大標本の標準体長は38cmで,他の種も30 cm前後です.最近のアンコウ目内の分子系統解析では,アンコウ科(=アンコウ亜目)の4属は単系統群であり,アンコウ目の中では最初に分岐するグループとされています(Miya et al., 2010).
参考文献
Eschmeyer, W.N. and J. D. Fong. 2011. Pisces. Z.-Q. Zhang, ed. in Animal biodiversity: an outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148: 26–38.
Ho, H.-C. and K.-T. Shao. 2008. A new species of anglerfish (Lophiidae: Lophiodes) from the western Pacific. Ichthyol. Res., 55: 367–373.
Ho, H.-C., B. Séret and K.-T. Shao. 2009. Redescription of Lophiodes infrabrunneus Smith and Radcliffe, 1912, a senior synonym of L. abdituspinus Ni, Wu and Li 1990 (Lophiiformes: Lophiidae). Zootaxa, 2326: 62–68.
Ho, H.-C., B. Séret and K.-T. Shao. 2011. Records of anglerfishes (Lophiiformes: Lophiidae) from the western South Pacific Ocean, with descriptions of two new species. J. Fish Biol., 79: 1722–1745.
Miya, M., T.W. Piersch, J.W. Orr, R.J. Arnold, T.P. Satoh, A.M. Shedlock, H.-C. Ho, M. Shimazaki, M. Yabe and M. Nishida. 2010. Evolutionary history of anglerfishes (Teleostei: Lophiiformes): a mitogenomic perspective. BMC Evolutionary Biology,10: 58
(1-27).
{kind=link}
岡村 収.1984.アンコウ科.岡村 収・北島忠弘(編),pp. 266-271, 376–378. 沖縄舟状海盆及び周辺海域の魚類 I.大陸棚斜面未利用資源精密調査.日本水産資源保護協会,東京.
写真標本データ:BSKU 93851,ca. 16 cm SL,2008年1月28日,高知県高知市御畳瀬魚市場(大手繰網),幸成丸,採集・写真:中山直英
(遠藤広光)
2011年11月の魚
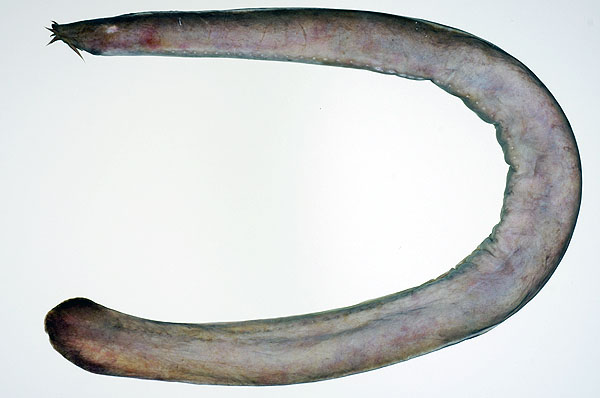
ヌタウナギ Eptatretus burgeri (Girard, 1855) (ヌタウナギ目ヌタウナギ科)
ヌタウナギは無顎上綱(Agnatha)ヌタウナギ綱に分類され,最初に魚類の系統から枝分かれしたグループと考えられています.この仲間には,現在1目1科2亜科5属76種が知られています(Nelson, 2006; Kuo et al., 2010; Knapp et al., 2011).そのうち,日本周辺には2亜科3属5種が分布します(岩田,2000).土佐湾では本種の他にムラサキヌタウナギ Eptatretus okinoseanus とクロヌタウナギ Eptatretus atami が記録されています.ヌタウナギ類は,少し前まで「メクラウナギ」の名称で広く知られていましたが,日本魚類学会の差別的標準和名の改称最終勧告(2007年)に従って,分類群の和名がメクラウナギからヌタウナギへと変更されています.
ヌタウナギ類は,体が細長くウナギ型で,顎がない,口の周辺には3~4対のひげをもち,吻端には外鼻孔が1つ開く,眼が退化的で皮下に埋没する,対鰭はなく,鰭は尾鰭のみで,体表に鱗はなく,皮膚は厚くなめらかで体側に並ぶ粘液線から大量の粘液を分泌する,硬骨組織がないなどの特徴をもちます(岩井,2005).顎はないのですが,口の中には2列に並んだ角質の鋭い歯列をもち,海底上にある魚やクジラの死体,漁網に掛かって弱った魚などの肉を削ぎ取って食べる腐食食者(スカベンジャー)とされてきました.しかし,小型の魚類や無脊椎動物も補食することが知られています(岩井,2005).ヌタウナギ属は,浅海から深海域まで広く生息し,本種は日本の本州中部以南から朝鮮半島南部,黄海と東シナ海に分布し,水深5~270 m から記録されています(岩田,2000; Knapp et al., 2011).また,食用として漁獲されることもあり,多くの海域で絶滅が危惧されています(Knapp et al., 2011).日本や韓国などでは開かずに2本の串に刺して蒲焼きにしますが,私はまだ食べたことがありません.
現世の無顎類はヌタウナギ類とヤツメウナギ類(頭甲綱ヤツメウナギ目ヤツメウナギ科)のみで,絶滅した無顎類には多様な化石種を含む甲皮類が知られています.顎をもつ現世の魚類は,両生類から哺乳類までを含む四足類とともに顎口上綱(Gnathostomata)に分類されています.無顎類は広義には魚類の仲間とされますが,ヌタウナギ類は脊椎骨をもっていないことから,ヌタウナギ類を含めたグループを有頭動物(Craniata)とし,ヌタウナギ類を除くヤツメウナギ類と顎口類のグループを脊椎動物(Vertebrata)とする見解もあります(詳しくは,中坊,2000).これはヌタウナギ類が最初に枝分かれした最も原始的な魚類で,無顎類が単系統群*とはならない仮説です(図1A).この仮説は,ヌタウナギ類が脊椎骨を欠くこと,リンパ形組織の特徴,脊椎動物の原始的な形態学的特徴を数多くもつことにより支持されてきました.しかし,最新の進化発生学分野の研究成果により,ヌタウナギは痕跡的な脊椎骨(軟骨)をもち,脊椎骨の形成に必要で脊椎動物の発生プログラムに用いられる遺伝子群が発現していることがわかりました(Ota et al., 2011)*下記のリンク参照.したがって,ヌタウナギ類の祖先は脊椎骨をすでに獲得していたと推定されるので,無顎類と顎口類をまとめて脊椎動物と呼んで間違いないことになります.もうひとつの仮説は,無顎類の2グループが単系統群となるものです(図1B).この仮説は近年のDNAやRNAを用いた分子系統解析により支持され,ヌタウナギ類が他の脊椎動物のもつ形態学的特徴を二次的に失ったか,それらが極めて退化しているという解釈になります.つまり,ヌタウナギ類がもつ特徴が,魚類の中では原始的なものなのか,あるいはそれらの多くは退化的であり(二次的に消失),原始的に見えるだけなのかわからないという点に原因がありました.最近,分子系統解析の情報の蓄積や形態学的な特徴の再検討の結果から,無顎類が単系統群*となる後者の仮説が有力のようです(Near, 2009; Heimberg et al., 2010; Ota et al., 2011).
*単系統群:1つの祖先から枝分かれしたグループ
をすべて含む
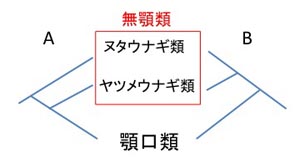
図1.現世の無顎類2グループと顎口類の系統仮説.
参考文献
Heimberg, A. M., R. Cowper-Sal・lari, M. Sémon, P. C. J. Donoghue and K. J. Peterson. 2010. microRNAs reveal the interrelationships of hagfish, lampreys, and gnathostomes and the nature of the ancestral vertebrate. PNAS, 107(45): 19379-19383.
岩井 保.2005.魚学入門.恒星社厚生閣,東京.220 pp.
岩田明久.2000.メクラウナギ科.中坊徹次 (編),pp. 107-109, 1439-1440. 日本産魚類検索 全種の同定.第2版.東海大学出版会,東京.
Knapp, L., M. M. Mincarone, H. Harwell, B. Polidoro, J. Sanciangco and K. Capenter. 2011. Conservation status of the world's hagfish species and the loss of phylogenetic diversity and ecosystem function. Aquatic Conserv.: Mar. Fresh. Ecosyst., 21: 401-411.
Kuo, C.-H., S.-C. Lee and H.-K. Mok. 2010. A new species of hagfish Eptatretus rubicundus (Myxinidae: Myxiniformes) from Taiwan, with reference to its phylognentic position based on its mitochondrial DNA sequence. Zool. Stud., 49(6): 855-864.
中坊徹次.2000.魚類概説 第二版.中坊徹次 (編),pp. xx-xliv. 日本産魚類検索 全種の同定.第2版.東海大学出版会,東京.
Near, T. J. 2009. Conflict and resolution between phylogenies inferred from molecular and phenotypic data sets for hagfish, lampreys, and gnathostomes. J. Exp. Zool. (Mol Dev Evol)312B: 749-761.
Nelson, J. S. 2006. Fishes of the world. Forth edition. John Wiley and Sons, Inc. Hoboken, New Jersey. 601pp.
Ota, K. G., S. Fujimoto, Y. Oishi and S. Kuratani. 2011. Identification of vertebra-like elements and their possible differentiation from sclerotomes in the hagfish. Nat. Commun. 2:373.doi:10.1038/ncomms1355 (2011)
*日本産魚類の差別的標準和名の改名最終勧告 *ヌタウナギの卵:写真1,写真2,写真3,写真4
*背骨を持たない脊椎動物「ヌタウナギ」に背骨の痕跡を発見(理化学研究所:プレスリリース 2011年6月29日)
写真標本データ:BSKU 92680,ca. 50 cm TL,2007年12月7日,高知県幡多郡黒潮町佐賀,佐賀漁港(底びき網),採集:太田真由加・坪井尚美,写真:中山直英
(遠藤広光)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2011年10月の魚

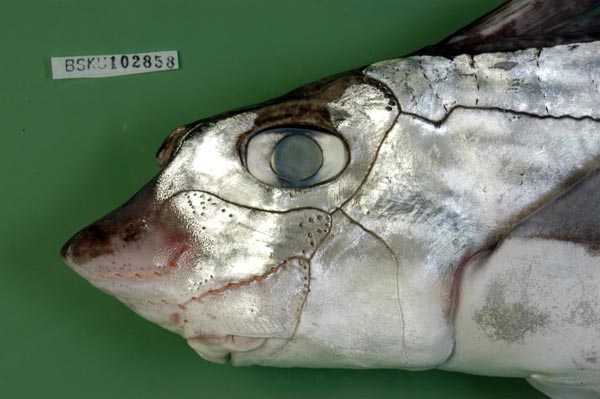
ギンザメ Chimaera phantasma Jordan et Snyder, 1900 (ギンザメ目ギンザメ科)
ギンザメ科 (Chimaeridae) は,北極と南極水域を除く世界中の海域に分布し, 沿岸域から水深2000 mを超える深海域まで生息する大型の軟骨魚類です (Didier et al., 2004). 本科はギンザメ属とアカギンザメ属の2属からなり,38有効種が知られています (Didier et al., 2004; Nelson, 2006; Kyne and Simpfendorfer, 2010). そのうち,日本周辺では2属8種が記録されています (Nakabo, 2002).成熟サイズは様々で,全長約60㎝の小型種から1mを超える大型種もいます.ギンザメ科魚類は,頭部が大きく,体が延長し,後方に向かって細くなっていきます.そして、本科の特徴である大きな胸鰭を上下に動かすことで,海中を羽ばたく様に優雅に泳ぎます.頭部の側線系はよく発達しており,まるで皮膚を繋ぎ合せたかのようです.本種は,生時には体が銀白色であること,体を走る側線が小刻みに波打つことで他の種と識別することができます.
高知県高知市の御畳瀬漁港の大手繰り網漁(沖合底曵き網)では,本種がよく水揚げされます.一度,お刺身が美味しいと聞いたので実際に試したところ,食べられるようなものではありませんでした.実際には練り製品に加工され市場に出回っているそうです.
{kind=link}
参考文献
Didier, D. A. 2004. Phylogeny and classification of extant Holocephalii, pp. 115-135. In J,C. Carrier, J.A. Musick and M. R. Heithaus (eds). Biology of sharks and their relatives. CRC Press, London.
Kyne, P. T. and C. A. Simpfendorfer. 2010. Deepwater Chondrichthyans. pp. 37-79. Sharks and their relatives Ⅱ. CRC Press, London.
中坊徹次.2000.ギンザメ科.中坊徹次 (編),pp. 113-114. 日本産魚類検索 全種の同定.第2版.東海大学出版会,東京.
Nelson, J. S. 2006. Fishes of the world. Forth edition. John Wiley and Sons, Inc. Hoboken, New Jersey. 601pp.
写真標本データ:BSKU 102858,828 mm TL,雄,2010年3月28日,高知市御畳瀬魚市場(大手繰り網),採集&写真:土居美月・中山直英
(土居美月)
2011年9月の魚

クロホシマンジュウダイ Scatophagus argus (Linnaeus, 1766) (スズキ目クロホシマンジュウダイ科)
クロホシマンジュウダイ科魚類は,インド・太平洋熱帯から亜熱帯沿岸域の内湾やマングローブに広く生息し,世界で2属4種のみが知られる小さなグループです.そのうち,日本にはクロホシマンジュウダイScatophagus argus (Linnaeus, 1766) のみが分布します(井田,1984).本種は,東南アジアでは観賞魚としてだけではなく,重要な食料資源とされています(Shao et al., 2004).また,本科のもう一方の属に分類されるSelenotoca multifasciata (Richardson,1864) は,形態的な特徴が本種とよく似る海外産の種で,シルバースキャットと呼ばれ,日本では観賞魚として人気があります.
日本では,本種はおもに琉球列島以南に分布し,幼魚は和歌山県や長崎県から報告があります(益田ほか,1975).幼魚は夏から秋にかけて河川や内湾の汽水や淡水域に出現し,しばらくは河口のセキショウモやオオカナダモが繁茂する場所で成育します.成魚はやや濁った内湾にすむという特徴があります(井田,1984).本種は幼魚期が過ぎると体に斑点が現れ,成長にともなって相対的に斑点のサイズは小さくなり,数は多くなります.成魚の体には40~50個の黒色斑点が散在し,体の地色は銀色を呈します(木下,1989).本科魚類の稚魚は,トリクチス期というチョウチョウウオ科の幼魚にも見られる幼生期を経ます.この時期の稚魚の頭部は,全体が骨板に覆われたような状態です.本種では体長25 mm まで,この特徴が残存します(木下,1989).
高知県では,本種は2002年に出版された「高知県レッドデータブック[動物編]」によって,準絶滅危惧種に指定されており,これまでに高知県からは四万十川(岡村・為家,1977;木下, 1989),柏島(平田ほか,1996),浦戸湾(町田・山川,2005),須崎湾(加藤ほか,2007)に生息することが報告されています.このうち,加藤ほか(2007)は須崎湾から採集された亜成魚を報告し,四万十川から浦戸湾に至る水域における出現を明らかにするとともに,高知県で再生産している可能性があるとしました.写真の個体は,浦戸湾の刺し網で得られた3個体の成魚のうちの1個体です.近年,高知県では各地で本種の成魚が発見されています.おそらく,これらの報告が示しているとおり,現在では高知県で繁殖をしている可能性が高いと考えられています(片山ほか,2009).
クロホシマンジュウダイはチョウチョウウオ科によく似た体型をしており,Linnaeus (1766)によりチョウチョウウオ属(Chaetodon) の種として記載されました.しかし,本種はチョウチョウウオ科の種に比べると,体は肉厚でやや厚いため,漁港などで釣りをしている人が,上から見て本種に気づくことがしばしばあるようです.現在,クロホシマンジュウダイ科はニザダイ亜目に分類され,系統学的にはスズキ亜目のチョウチョウウオ科と近縁ではないと考えられています.図鑑類では,本科はアイゴ科の直前に出てきます(例えば,Nakabo, 2002; Nelson, 2006).また,本科は背鰭や臀鰭の棘に毒腺をもち,弱毒を分泌することが知られていますが(Cameron and Endean, 1970),硬骨魚類の毒腺の進化に関する最近の系統仮説では,同様の毒腺をもつアイゴ科との近縁性は示されていません (Smith and Wheeler, 2006).一方,最近の核のDNAに基づくニザダイ上科内の分子系統解析では,本科はアイゴ科の姉妹群とされています(Holcroft and Wiley, 2008).
{kind=link}
参考文献
Cameron, A. M. and R. Endean. 1970. Venom glands in scatophagid fish. Toxicon, 8(2): 171-174.
平田智法・山川武・岩田明久・真鍋三郎・平松 亘・大西信弘.1996.高知県柏島の魚類相-行動と生態に関する記述を中心として-.高知大学海洋生物教育研究センター研究報告,(16)1–177.
Holocroft, N. I. and E. O. Wiley. 2008. Acanthuroid relationships revisited: a new nuclear gene-based analysis that incorporates tetraodontiform representatives. Ichthyol. Res., 55: 274-283.
井田 齊.1984.クロホシマンジュウダイ科.(益田一・尼岡邦夫・荒賀忠一・上野輝彌・吉野哲夫,編)pp. 176.日本産魚類大図鑑.東海大学出版会,東京.
片山英里・阪本匡祥・渡辺博満・中村和喜・町田吉彦.2009. 高知県浦戸湾で得られたクロホシマンジュウダイの成魚と香南市香宗川で得られた幼魚(スズキ目クロホシマンジュウダイ科).四国自然史科学研究,(5): 11-14.
加藤正洋・石川晃寛・伊佐正樹・町田吉彦.2007.クロホシマンジュウダイの須崎湾からの初記録.四国自然史科学研究, (4)24-26.
木下 泉.1989.クロホシマンジュウダイ.川那辺浩哉・水野信彦(編)日本の淡水魚.p. 533.山と渓谷社,東京.
高知県レッドデータブック[動物編]編集委員会(編).2002.高知県レッドデータブック[動物編].高知県文化環境部環境保全課,高知. 470pp.
町田吉彦・山川武.2005.浦戸湾初記録を含む高知県産クロホシマンジュウダイの標本(スズキ目クロホシマンジュウダイ科).四国自然史科学研究,(2)58-62.
益田 一・荒賀忠一・吉野哲夫.1975.魚類図鑑 南日本の沿岸魚.東海大学出版会,東京,379 pp.
Nakabo, T., ed. 2002.
Fishes of Japan with pictorial keys to the species, English edition. Tokai University Press, Tokyo. i-lxi, 1749 pp.
Nelson, J. S. 2006. Fishes of the world. 4th ed. John Wiley & Sons, Hoboken. 601pp.
岡村 収・為家節弥.1977.四万十川の魚類(高知県編:四万十川水系の生物と環境に関する総合調査-1976年度委託調査-),高知,159¬−232 pp.
Shao, Y.-T., L.-Y. Hwang and T.-H. Lee. 2004. Histological observations of ovotestis in the spotted scat Scatophagus argus.
Fish. Sci. 70: 716–718.
Smith, W. L. and W. C. Wheeler. 2006. Venom evolution widespread in fishes: a phylogenetic road map for the bioprospecting of piscine venoms. J. Heredity, 97(3): 206-217.
写真標本データ:BSKU 92385,252.5 mm SL,高知県高知市浦戸湾,刺し網, 2007年11月21日;BSKU 93510,25.0 mm SL,幼魚,高知県香南市香宗川,手網,セキショウモ群落,採集:中村和喜,2007年9月30日.
(片山英里・遠藤広光)
2011年8月の魚

ズグロダテハゼ Amblyeleotris melanocephala Aonuma, Iwata et Yoshino, 2000 (スズキ目ハゼ亜目ハゼ科)
ダテハゼ属はインド−西太平洋から約33種が知られ,日本には13種ほどが分布するとされています(鈴木ほか,2006).ダテハゼ属魚類は岩礁やサンゴ礁域の砂底に生息し,テッポウエビ類と共生することがよく知られています.巣穴の近くで雌雄がかわいらしく並んでいる姿が見られるため,ダイバーにも人気の高いグループの1つです.しかし,テッポウエビ類と共生するハゼ類は,いずれも警戒心が強く,危険を察知するとすぐに巣穴に入ってしまいます.高知県では,平田ほか(1996)が柏島から本属7 種を報告しており,本研究室の最近の調査も加えると9種の分布が確認されています.本属は未記載種や初記録種が含まれ,現在でも分類学的研究が進められています.最近では,2007年にモリシタダテハゼが新種記載されています(Senou and Aonuma, 2007).
ズグロダテハゼは,2000年に高知県と沖縄県から得られた標本に基づいて記載されました(Aonuma et al., 2000).ホロタイプは高知県最南西部に位置する沖の島の久保浦の水深28 m から採集されています.2010年にはホロタイプの採集場所とほぼ同じ,久保浦の水深30mから2個体が採集されました.そのうちの1個体が今回の標本写真です.本種は水深30~50m付近のやや深場に生息し,高知県南西部以南では普通にみられるようです.頭は吻から鰓蓋にかけて黒色を帯びて,体には4本の黄褐色黄帯があることが特徴です.日本産の本属の中で,このように頭が黒くなるのは本種のみです.また,本属の腹鰭は吸盤状ですが,本種ではU字状の切れ込みがあります.
本種の和名は見た目どおりの「ズグロ」で,学名のうち“melanocephala”という種小名もまたギリシャ語で頭が黒いという意味です.水中では胸鰭基底に黄色斑点があり,背鰭の朱色の縁取りも鮮やかで,他のダテハゼ類と同様にかわいらしい姿をしています.しかし,標本にするととても貫禄のある顔になります.
参考文献
Aonuma, Y., A. Iwata and T. Yoshino. 2000. A new species of the genus Amblyeleotris (Pisces: Gobiidae) from Japan. Ichthyol. Res., 47(2): 113–117.
平田智法・山川 武・岩田明久・真鍋三郎・平松 亘・大西信弘.1996.高知県柏島の魚類相.行動と生態に関する記述を中心として.Bull. Mar. Sci. Fish., Kochi Univ., 16: 1-177.
Senou, H. and Y. Aonuma, 2007. A new shrimp goby of the genus Amblyeleotris (Perciformes: Gobiidae) from the Ogasawara Islands, Japan. Bull. Natl. Mus. Nat. Sci., Ser. A, Suppl. 1: 101-107.
鈴木寿之・渋川浩一・矢野維幾(瀬能 宏,監修).2004.決定版 日本のハゼ.536pp.
写真標本データ:BSKU 103945, 79 mm SL,2010年7月21日,高知県宿毛市沖の島久保浦,水深30m,採集:坪井美由紀,写真撮影:遠藤広光 白バックの写真
(片山英里)
{kind=link}
2011年7月の魚

Suttonia sp. (スズキ目ハタ科ハタ亜科ヌノサラシ族)
ハタ科魚類は世界中で64属475種が知られ,熱帯から温帯浅海域の岩礁やサンゴ礁から沖合の大陸棚まで広く生息しています.本科には最大体長 3 m,重量 400 kg に達する種もいますが,多くは10cmよりも大きくなりません(Nelson,2006).
Suttonia 属は,熱帯域から亜熱帯域に生息する浅海性の小型のハタ亜科ヌノサラシ族魚類です.西部インド洋に分布する S. suttoni と東部インド洋から中部太平洋にかけて広く分布する S. lineata の2種が知られており,日本周辺では本属の生息が確認されていませんでした.本属は体色が一様に赤色で,下顎の先端から背鰭起部までの正中線上に薄桃色の縦帯をもつ,円鱗をもつ,サーベル状の大きな前鰓蓋骨棘をもつ,前鼻管が長いなどの特徴により,他の本科の属とは容易に識別できます.また,Suttonia 属も他のヌノサラシ族のいくつかの種と同様に,体表から粘液毒のグラスミチンを分泌することが知られています(Randall and Baldwin, 1997).
2010年7月に行った高知県宿毛市沖の島沿岸の潜水調査で,久保浦港近くの水深16mにある岩の下から,本属の1標本(写真個体)を採集しました.この標本は,尾柄長が短い,縦列鱗数51,脊椎骨数10+16などの特徴から S. suttoni に似ていますが,背鰭と臀鰭の最大棘長が短い,有孔側線鱗数26,胸鰭の腋部が被鱗する,頭部背面が暗褐色であるなどで明瞭に異なります.したがって,写真の標本は,本属の未記載種の可能性があります.
参考文献
Nelson, J. S. 2006. Fishes of the world. 4th ed. John Wiley & Sons, Hoboken. 601pp.
Randall, J. E. and C. C. Baldwin. 1997. Revision of the serranid fishes of the subtribe Pseudogrammina, with descriptions of five new species. Indo-Pacific Fishes, (26): 1–56.
写真標本データ:BSKU 103900, 65 mm SL,2010年7月21日,高知県宿毛市沖の島久保浦,水深16m,採集・写真撮影:遠藤広光
(劔物健太・遠藤広光)
2011年6月の魚
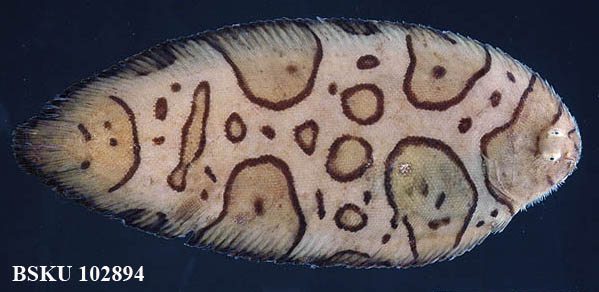
ワモンウシノシタ Brachirus annularis Fowler, 1934(カレイ目ササウシノシタ科)
ササウシノシタ科のワモンウシノシタは,1993年3月に土佐湾中央部での豊旗丸のトロール調査で採集された1標本に基づいて,日本初記録として報告されました(Gonzales et al., 1994).本種は日本では土佐湾のみで記録され,国外では台湾,フィリピン(基産地),そしてオーストラリアに分布しています.土佐湾で最初の標本が採集された翌年の3月と昨年の4月には,高知市の御畳瀬魚市場(いずれも幸成丸の大手繰り網)で,それぞれ1個体が採集されています.有眼側の体の縁辺から大きく広がる斑紋は,頭部と尾部,その間に2対の合計6つあることが本種の特徴ですが,土佐湾産の3標本を比較すると,焦げ茶色の縁取りのある大小の輪状斑の形には個体変異が見られます.
ワモンウシノシタの学名の種小名"annularis"は「環状の,輪状の」という意味で,和名「ワモン」と同様にその特徴的な有眼側の模様に因んでいます.本種は Brachirus 属として Fowler (1934)により記載され,その後 ミナミシマウシノシタ属(Synaptura )へ移されていました(中坊,2000).Desoutter et al. (2001)はササウシノシタ科の3属BrachirusとSynaptura ,そして Euryglossa についての有効性を検討し,Brachirus を有効属として,Synaptura はそのシノニム(異名)と判断しました.Swainson (1839)は Brachirus をササウシノシタ科の新属とした論文の中で,綴りが1文字違いのフサカサゴ科の新亜属 Brachyrus も設立しています.しかし,論文中の記載以外の部分で,フサカサゴ科の Brachyrus とすべきところを,"Brachirus" と綴り間違いをしたことから,Brachyrus のホモニム(同名)との見解もあり,無効と見なされていました.属階級群名については,ひと文字違いは別の名前とみなされ,ササウシノシタ科内の属名との検討からも有効な属とされています(中坊,2000; Nakabo, 2002).
Gonzales et al. (1994)の第1著者のBenjamin J. Gonzalesさんはフィリピンからの留学生で,私たちの研究室でネズッポ科魚類の生態学的研究を行い,1994年3月に修士の学位を取得しています.ワモンウシノシタが豊旗丸で採集された時には,乗船していたかもしれません.
参考文献
Desoutter, M., T.A. Munroe and F. Chapleau. 2001. Nomenclatural status of Brachirus Swainson, Synaptura Cantor and Euryglossa Kaup (Soleidae, Pleuronectiformes). Ichthyol. Res., 48(3): 325–327.
Fowler, H.W. 1934. Descriptions of new fishes obtained 1907 to 1910, chiefly in the Philippine Islands and adjacent seas. Proc. Acad. Nat. Sci. Philad., 85: 233-367.
Gonzales, B.J., O. Okamura, K. Nakamura and H. Miyahara. 1994. New record of the annular sole, Synaptura annularis (Soleidae, Pleuronectiformes) from Japan. Japan. J. Ichthyol., 40(4): 491–494.
中坊徹次.2000.ササウシノシタ科.pp.1383-1387, 1638-1639.中坊徹次,編.日本産魚類検索 全種の同定.第2版.東海大学出版会,東京.
Nakabo, T. 2002.
Soleidae. pp. 1383-1387, 1629-1630. T. Nakabo, ed. Fishes of Japan with pictorial keys to the species, English edition. Tokai University Press, Tokyo.
*本種は2009年11月に三重県尾鷲市でも採集されています【三日に一魚の尾鷲市役所のページ】.
写真標本データ:BSKU 102894, 155 mm SL,2010年4月11日 土佐湾,高知市御畳瀬魚市場大手繰り網,幸成丸,採集・写真撮影:土居美月
他の標本:BSKU 81384, 122 mm SL, 土佐湾中央部,水深151–154m,豊旗丸,ビームトロール,1993年3月5日(Gonzales et al., 1994で報告); BSKU 81717, 113 mm SL, 土佐湾中央部,水深約180m,高知市御畳瀬魚市場、幸成丸,1994年3月16日.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(遠藤広光)
2011年5月の魚

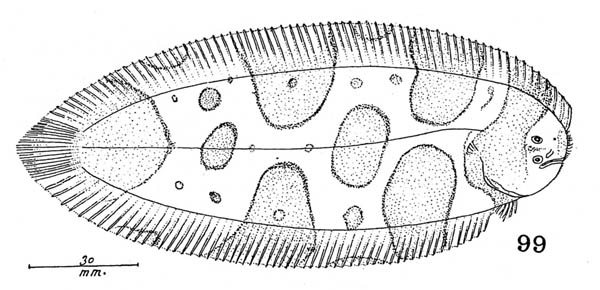
ベニテグリ Foetorepus altivelis (Temminck et Schlegel, 1845)(スズキ目ネズッポ科)
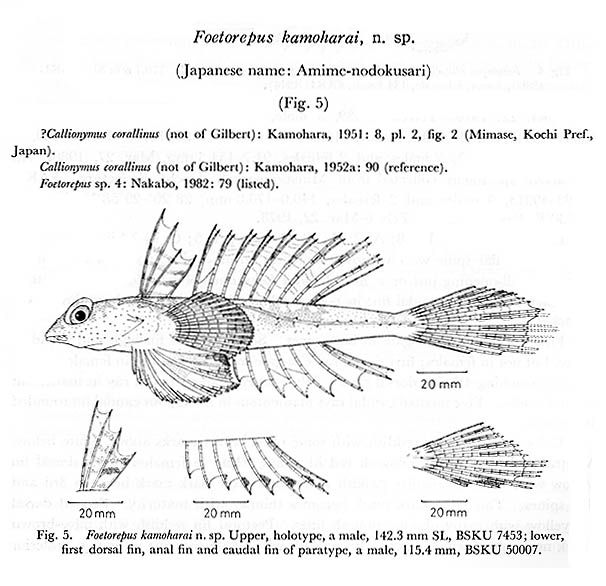
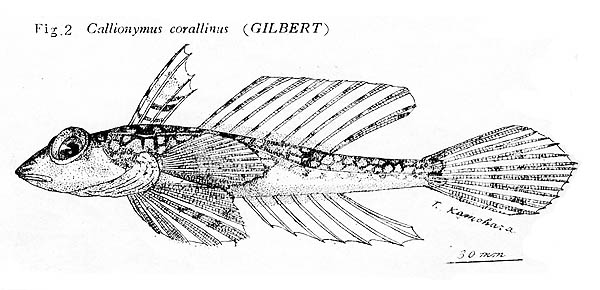
ベニテグリは南日本の太平洋岸から東シナ海,そして南シナ海北部の大陸棚縁辺域に生息するネズッポ科魚類です.日本産のベニテグリ属はベニテグリFoetorepus altivelis,アミメノドクサリFoetorepus kamoharai Nakabo, 1983, そしてルソンベニテグリFoetorepus masudai Nakabo, 1987の3種です.ベニテグリ属はその名の通り朱色の体が特徴で,第2背鰭には黄色や桃色,白色など種ごとに異なる模様をもちとても鮮やかです.高知県の御畳瀬漁港ではベニテグリとルソンベニテグリの2種がよく見られ,標本数も豊富です.この2種は第1背鰭と第2背鰭の斑紋などで,容易に区別することができます.同属のアミメノドクサリは,体側後半部の側線から出る短枝が上下へ向くことや第1背鰭に4本の暗色線があることで,他の2種と区別することができます.しかし,アミメノドクサリは,基産地(タイプ標本の産地)の高知県からも他の海域からも追加記録のない稀種です.
ベニテグリはふだん店頭で見かける魚ではありませんが,皮を1度で剥ぐことができ,身がしっかりしていて食用になります.いろいろと試してみた結果,刺身,塩焼き,天ぷら,とくに干物にすると美味しく頂けます.
アミメノドクサリの種小名 "kamoharai"は,本研究室の初代教授である蒲原稔治博士に献名されたものです.京都大学の中坊徹次博士は,本研究室に所蔵されていた高知県産(御畳瀬魚市場で採集)の2標本をホロタイプ(BSKU 7452, 142.3 mm SL,1956年10月5日)とパラタイプ(BSKU 50007, 115.4 mm SL,1965年2月20日) として本種を記載しました.これは蒲原博士が1951年の論文で,御畳瀬魚市場で採集された本種の1標本(ca. 165 mm SL[図からの推定], 220 mm TL)をハワイ諸島から知られる ?Callionymus corallinus Gilbert, 1905 アミメノドクサリ(New Japanese name)として報告していたためです.Nakabo (1983)はKamohara (1951)の標本が本種のホロタイプであるとしましたが,ホロタイプは1956年に採集されたので,蒲原博士が最初に報告した標本は1951年以前に採集された別のものであることは確かです.そうなると,Kamohara (1951)の標本が本研究室の標本コレクションの中にあるのか,行方不明になってしまったのか気になります.BSKU標本台帳の 7452には "Synchiropus calauropomus "と書かれていますが,この種はオーストラリアやニュージーランド周辺に分布する Foetorepus calauropomus (Richardson, 1844)で,日本周辺には分布しません.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
参考文献
Kamohara, T. 1951. Notes on some rare fishes from Prov. Tosa, Japan. Rep. Kochi Univ., Nat. Sci., (1): 1-8, pls 2.
Kamohara, T. 1952. Revised descriptions of the offshore bottom-fishes of Prov. Tosa, Shikoku, Japan. Rep. Kochi Univ., Nat. Sci., (3): 1-122.
Nakabo, T. 1983. Revision of the dragonets (Pisces: Callionymidae) found in the waters of Japan. Publ. Seto Mar. Biol. Lab., 27 (4/6): 193-259.
Nakabo, T. 1987. A new species of the genus Foetorepus (Callionymidae) from southern Japan with a revised key to the Japanese species of the genus. Japan. J. Ichthyol., 33(4): 335–341.
中坊徹次.2000.ネズッポ科.中坊徹次,(編)pp.1125-1138.日本産魚類検索 全種の同定.第2版.東海大学出版会,東京.
標本写真データ
BSKU 94908, 84.8 mm SL, 土佐湾中央部,水深150m,中央水産研究所調査船こたか丸,St.3-2,オッタートロール,2008年5月22日,採集・写真撮影:中山直英.
(廣田愛実・遠藤広光)
2011年4月の魚

タイワンコロザメ Squatina formosa Shen et Ting, 1972(カスザメ目カスザメ科)
(頭部,腹面,尾部の写真)
カスザメ属(genus Squatina)は,最大で2メートルに達する底生性のサメ類で,大西洋,インド洋南部,そして太平洋の熱帯・亜熱帯水域に分布し,沿岸から大陸棚斜面上部にかけて生息します(Nelson, 2006).カスザメ目カスザメ科に含まれる唯一のグループで,現在世界から18種が知られています(Last and White, 2008).本属は本属は頭と体は扁平で幅広く,胸鰭と腹鰭も大きく,全体的にやや菱形をしています.一方,尾部はやや長く,断面が丸いので,サメとエイの中間的な体型に見えます.普段はこの扁平な体で砂地に潜んでおり,餌となる魚類が頭の上を通り過ぎる瞬間に頭を持ち上げて丸呑みにします.本属魚類の英名は“Angel shark”ですが,その名前には似つかわしくない摂餌行動といえるでしょう.
日本周辺にはカスザメ S. japonica (Bleeker, 1858),コロザメ S. nebulosa (Regan, 1906)とタイワンコロザメ S. formosa Shen and Ting, 1972 の3種が分布しています(Kriwet et al., 2010).北西太平洋域には,これら3種と S. tergocellatoides (Chen, 1963) [南シナ海から台湾]の4種が分布し,Walsh and Ebert (2008)により分類的に再検討されています.カスザメとS. tergocellatoides は,腹鰭先端が第1背鰭の基底に達しないことで,他の2種と異なります.また,カスザメは体の背側正中線上に棘列をもちますが,他の3種にはありません.一方,コロザメとタイワンコロザメは特徴がよく似ており,これまでも混同されてきました.とくに日本産の本属は,背側に棘列をもつカスザメともたないコロザメの2種とされてきたため,疑いもなくタイワンコロザメも“コロザメ”に同定されていました.2008年にドイツ・ベルリンにあるフンボルト大学の大学院生であったStelbrinkさんにコロザメの分子解析用サンプルを頼まれて(Stelbrink et al., 2010),土佐湾のコロザメと思って送ったものが台湾周辺でのみ記録のある別種 S. formosa と判明して大変驚きました.そのような経緯から,Stelbrinkさんと彼の先生であったKriwetさんと共著で日本初記録の論文を出しました(Kriwet et al., 2010).種小名の“formosa”は台湾を意味するので,この論文の中で“タイワンコロザメ”という新標準和名を提唱しています.
Walsh and Ebert (2008)によると,コロザメとタイワンコロザメは,上唇付近の形,背鰭や尾鰭の形(コロザメでは縁辺が角張っているが,タイワンコロザメでは丸みを帯びる)や腰骨間の幅に違いがあります.また,タイワンコロザメは2つの背鰭下方の体側に大きな円形の褐色斑紋もちますが,コロザメでは小斑点であり,あまり目立ちません.さらに,タイワンコロザメではコロザメに比べて,体の背面で左右対称に位置する小白色斑点はよく目立ちます.Walsh and Ebert (2008)はこれらの斑紋や斑点を両種の識別形質としていないので,成長や個体による変異について,もう少し調べる必要がありそうです.また,両種の日本沿岸における分布も,標本に基づいて調べ直さなければなりません.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
参考文献
Kriwet, J., H. Endo and B. Stelbrink. 2010. On the occurence of the Tiwan angel shark, Squatina formosa Shen & Ting, 1972 (Chondrichthyes, Squatinidae) from Japan. Zoosyst. Evol., 86(1): 117–124.
Last, P. R. and W. T. White. 2008. Three new angel sharks (Chondrichthyes: Squatinidae) from the Indo-Australian region. Zootaxa, 1734: 1–26.
Nelson, J. S. 2006. Fishes of the world. 4h ed. John Wiley and Sons, Inc.. Hoboken, New Jersey. 601pp.
Stelbrink, B., T. von Rintelen, G. Cliff and J. Kriwet. 2010. Molecular systematics and global phylogeography of angel sharks (genus Squatina). Molecular Phylogenetics and Evolution, 54: 395–404.
Walsh, J. and D. A. Ebert. 2007. A review of the systematics of western North Pacific angel sharks, genus Squatina, with redescriptions of Squatina formosa, S. japonica, and S. nebulosa (Chondrichthyes: Squatiniformes, Squatinidae). Zootaxa, 1551: 31–47.
標本写真データ
BSKU 95231,546 mm TL,2008年6月16日,土佐湾中央部 水深約200 m,中央水産研究所調査船こたか丸,オッタートロール,採集:中山直英,写真撮影: 遠藤広光.
BSKU29904, 1077 mm TL, 1979年11月4日,沖縄舟状海盆,水深300–330m.
(遠藤広光)
2011年3月の魚

ホソウケグチヒイラギ Secutor indicius Monkolprasit, 1973(スズキ目ヒイラギ科)写真2
{kind=link}
ヒイラギ科の魚類は主に内湾や沿岸の浅所および河口から河川汽水域の砂泥底に生息しており(木村,2007),日本では5属13種が知られています(木村ほか,2008).ホソウケグチヒイラギはウケグチヒイラギ属に属しており,日本においては本種とヤンバルウケグチヒイラギの2種が知られています.本属の特徴は口が上向きで前上方に伸びること,口は閉じた状態だとほぼ垂直になることなどで本科他属と区別することができます(木村ほか,2008).
ホソウケグチヒイラギは鹿児島県内之浦湾で採集された標本に基づき,日本初記録として報告されました(木村ほか,2008).本種は体高が低いこと,体側にある横縞がほぼ垂直で15 – 22本あることなどで同属他種と見分けることができ,和名の「ホソ」は,この体高が低い(体が他種に比べやや細長い)という特徴に因んで付けられています(木村ほか,2008).
ヒイラギはサビキ釣りの対象魚として知られており,高知県ではニロギと呼ばれています.数釣りが楽しめ,食味もよく人気のある魚です.写真の標本も地元の方が香南市吉川漁港にて釣りで採集されたものです.本種は高知県沿岸で発見の報告がなく初記録であり,これまで知られていた分布の北限記録です.地元の方の御協力がなければ,今後も発見はされていなかったかもしれません.もし,釣りで見慣れない魚が釣れたときは逃がしてしまうのではなく,どんな種名の魚か調べてみるのも良いかもしれません.それが新たな発見につながる可能性があります.
参考文献
木村清志・伯耆匠二・山寺守彦・本村浩之.2008.鹿児島県で採集された日本初記録のヒイラギ科魚類ホソウケグチヒイラギ(新称)Secutor indicius .魚類学雑誌,55(2): 111 – 114.
木村清志・木村良子・池島 耕・本村浩之・岩槻幸雄・吉野哲夫.2008.ヒイラギ科魚類各属の標準和名.魚類学雑誌,55(1):62 – 63.
木村清志.2007.ヒイラギ科.岡村 収・尼岡邦夫編.山溪カラー名鑑:日本の海水魚.山と渓谷社,東京,pp.327.
標本写真データ
BSKU104501,94.9 mm SL,2010年10月22日,高知県香南市吉川漁港,採集・寄贈:竹内健二,写真撮影 劔物健太.
(劔物健太)
2011年2月の魚

セダカギンポ Exallias brevis (Kner, 1868)(スズキ目イソギンポ科)
イソギンポ科はイソギンポ族,カエルウオ族,ナベカ族およびニジギンポ族という4つの族(tribe)からなり,族によって形態や生態は様々です.世界中の熱帯・温帯域に分布し,53属約300種,日本でも28属約69種が知られ,岩礁域やサンゴ礁域,タイドプールに生息しています.体長は大きくても10cmほどにしかなりません(藍澤,2009).
セダカギンポはカエルウオ族セダカギンポ属に属し,世界ではインド洋と西部太平洋の熱帯域を中心に,国内では紀伊半島以南の太平洋岸,小笠原諸島と琉球列島に分布します(藍澤,2000).水深10m以浅の波当たりの強い岩礁性海岸やサンゴ礁に生息しており,岩礁などに付着した珪藻などをはみ取って食べています(藍澤,2000).本種の体色は雌雄差が明瞭で,雄では赤みを帯び,雌では比較的地味な色合いです.また,セダカギンポを含むイソギンポ科では,雄がもつ巣に複数の雌が訪れて産卵する“なわばり型複婚”という繁殖様式をとります(宮野ほか,2006).
本種の特徴はずっと見ていると目がチカチカするような網目斑紋です.また,「セダカ」の名前の通り,体高が他のギンポ亜目の種に比べると高く,ずんぐりとして,ふてぶてしくも憎めない顔つきをしています.そのためダイバーからの人気も高いですが,造礁サンゴ類の枝の隙間に隠れているため,なかなかお目にかかれないようです.
この個体は2010年7月に高知県宿毛市の沖の島でおこなわれた調査で採集されたものです.沖の島調査は研究室の毎年恒例の行事となっており,日中は採集,夜は標本作成や写真撮影を行います.3日間休む時間もほとんど無く過ごすので,最後は気力で持ちこたえます.その甲斐あってか,この調査によって高知県初記録種や稀少種が採集されています.重労働ですが,得るものは多いのでぜひ次回も参加したいと思います.
参考文献
藍澤正宏.2000.イソギンポ科.中坊徹次,(編)pp.1090-1119.日本産魚類検索 全 種の同定.第2版.東海大学出版会,東京.
藍澤正宏.2009.イソギンポ科.岡村収・尼岡邦夫,(編)pp.562-571.山渓カラー名 鑑 日本の海水魚.第3版.山と渓谷社,東京.
桑村 哲生.1996.魚類の繁殖戦略入門.桑村哲生・中嶋康裕,(編)pp.1-41.魚類の 繁殖行動Ⅰ.海游社,東京
宮野哲平・竹垣毅・夏刈豊.2006.ロウソクギンポの産卵・卵保護行動.長崎大学水産学部研 究報告,87:1-5.
標本写真データ
BSKU103990,66.6 mm SL,2010年 7月22日,高知県宿毛市沖の島,写真撮影:遠藤広光
(木村祐貴)
2011年1月の魚

オオミミズハゼ Luciogobius grandis Arai, 1970(ハゼ目ハゼ科)
ミミズハゼ属は体長5 cm前後の小型のハゼ科魚類で,ロシア,中国,朝鮮,日本にかけての極東アジアのみに分布しています.現在は世界で約16種,そのうち日本では約14種が知られています.本属魚類の特徴は,ミミズのような長い円筒形の体と,背鰭が体の後半部に1つのみであり,扁平した頭部をもちます.本属は種によって多少異なりますが,純淡水から汽水,海水と幅広い生息域で見られますが,礫間を好むため,あまり人目にはつきません.
オオミミズハゼはTomiyama (1936) によりミミズハゼ (L. guttatus Gill, 1859)の4型のうちの一つとして報告され,後にArai (1970) により新種として発表されました.体は無鱗で,眼は皮下に埋没せず,体長は本属内では最も大きく最大で約10 cmとなり,胸鰭の上方と下方に数本の遊離軟条があることから近縁種と容易に識別できます.また,本種は外海に面した海岸の転石の間隙に生息します(藍澤, 1997).
近縁種であるイドミミズハゼ(L. pallidus Regan, 1940)とドウクツミミズハゼ(L. albus Regan, 1940)は,それぞれ絶滅危惧IB類と情報不足とされました(高知県RDB).また,本属魚類は分類上の問題点を多く含み,現在ミミズハゼとされている種の中に十数種の未記載種が含まれると予想されています(鈴木他,2004).
本属は礫間にイカの足を垂らすと,釣り針無しでも容易に釣れるそうです.かき揚げにすると美味しく頂けるそうですが,釣りの餌として使われます.
参考文献
Arai, R., 1970. Luciogobius grandis, a new goby from Japan and Korea. Bull. Natn. Sci. Mus., Tokyo, 13(2):199-206, pl. 1.
藍澤 正宏.2004.オオミミズハゼ.p. 620 in 岡村 収・尼岡邦夫,編.日本の海水魚 第3版,山と渓谷社,東京.
高知県レッドデータブック〔動物編〕編集委員会(編).2002.高知県レッドデータブック〔動物編〕.高知県文化環境部環境保全課,高知,470pp.
鈴木寿之・渋川浩一・矢野維幾(瀬能 宏,監修).2004.決定版 日本のハゼ.536pp.
写真標本データ
BSKU 95264,54.4 mm SL 2008年7月7日,高知県香南市手結海岸,採集・写真撮影 朝田貴之
(坪井尚美)