2025�N12���̋�
 �@
�@
�c���O�G Liopropoma latifasciatnm (Tanaka, 1922)�i�X�Y�L�ڃn�i�X�Y�L�ȁj
�@�c���O�G Liopropoma latifasciatnm (Tanaka, 1922)�̓X�Y�L�ڃn�^�Ȃ��邢�̓n�i�X�Y�L�Ȃ̃n�i�X�Y�L���ɕ��ނ���C����⏬�����C���Z�����ЁC�̑��ɂ͂P�{�̍��F�c�т��݂��C�w�h��10��13����C�\�h��3��9����C���h��14-16����i�ʏ�15����j�C����ё����L���ؐ���47-52�Ƃ��������������܂��i�Ⴆ�CKatayama,1960�GRandall and Taylor, 1988�G���\�C2013�G���C2025�Gvan der Laan et al., 2025�j�D�n�i�X�Y�L���̋��ނ͐��E��40���`��33�L���킪�m���C���̂������{�ɂ̓c���O�G���܂߂�12�킪�������Ă��܂��iFricke et al, 2025�G�{���C2025�j�D�{���̓n�^�ނ̒��ł͑̃T�C�Y����r�I���^�ŁC�ŏ��̎�ł͑̒�70 mm�C�ő�̎�ł͑̒� 220 mm �Ƒ傫���ɕ�������܂��iRandall and Taylor, 1988; Koeda et al., 2019�j�D���̂����C�c���O�G�͍ő�̒��� 160 mm �ɒB���邽�߁C�{���ł͒��^��Ƃ����܂��iRandall and Taylor, 1988�G�r�c�E���V�C2015�j�D
�@�c���O�G�͓����鍑��w���w�����c���Ε䔎�m �i1878-1974�j�ɂ��C�����ߍx�i�����s��j�Ƙa�̎R���I�ɓc�Ӂi�c�ӎs���j�œ���ꂽ�W�{�Ɋ�Â��CPikea latifasciata�Ƃ��ĐV��L�ڂ���܂����i�c��, 1922�GRandall and Taylor, 1988�j�DCatalog of fishes�Ⓦ����w�̃f�[�^�x�[�X�ɂ��ƁC�z���^�C�v�iZUMT 8000�C�a�̎R���c�ӎs���C�I�ɐ����j�ƃp���^�C�v�iZUMT 3423�C�����s��j�̕W�{�́C������w�������������ق̋��ޕW�{�R���N�V�����ɕۊǂ���Ă��܂��iFricke et al., 2025�G������w�C2025�j�{��͋߉���̃N���I�r�X�Y�L L. lemniscatum Randall and Taylor, 1988�ɗގ����܂����C�̑��̍��F�c�т�������ł͑������痣��C�w�h��𐔂��ʏ�13������\�h��𐔂�9�ł��邱�Ƃ��环�ʂł��܂��i�Ⴆ�C���\�C2013�G�k�E�����C2025�G���C2025�j�D
�@�n�i�X�Y�L���̊w�� Liopropoma �́C�M���V����Łu���炩�C���R�ȁv�̈Ӗ��́uleios�v�ƁC�M���V����ł��ꂼ��u�`�̋߂��Ɂv�́upros�v����сu�W�C�����v�́upoma�v�̕�����ł��i���V�E�����C2015�j�D����C�c���O�G�̎포�� latifasciatnm �̓��e����Łu���L���т̂���v�Ƃ����Ӗ��������C���̐��ɍ��킹�Č���𒆐��`�ɕς������̂ŁC�{��̑傫�ȓ�����1�ł��鍕�F�c�т����O�̗R���ƍl�����܂��i���V�E�����C2015�j�D
�@�{��͐��[ 30 m ���� 220 m �̉��ݕ���嗤�I�Ζʂ̊�ʈ�ɒP�ƂŐ������C���ɐ��[ 30 m �Ȑ[�̊R�̌��ȂǂŊώ@����邱�Ƃ������C����{�̑����m�݂�ɓ��哇�C�����C�C�O�ł͊؍����p�C�p���I��������m���Ă��܂��i�g��E���\�C2008�C2023�G�r�c�E���V�C2015�j�D�X�N�[�o�_�C�r���O�Ńc���O�G�����ڋ߂���ƁC���A�g���B�����������邻���ł��i�g��E���\�C2008�C2023�j�D�v�c�ق��i1975�j�ɂ��ƁC�{��͌̐��I�ɂ݂Ă����Ȃ����ł��邽�߁C�C���ŏo�����ƂĂ����b�L�[�ł��ˁD
�Q�l����
Fricke, R., W. N. Eschmeyer and R. Van der Laan (eds.) 2025. Eschmeyer�fs catalog of fishes: genera, species, references. http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp Accessed 30 Nov. 2025.
�r�c�����E���V�O���D2015�D����{�����m���݂̋��ށD���C��w�o�ŕ��C�`��D597 pp.
Katayama, M. 1960. Fauna Japonica Serranidae (Pisces). Biogeographical Society of Japan, Tokyo. i-viii + 1-189 pp., pls. 1-86.
�v�c ��E�r�ꒉ��E�g��N�v�D1975�D���ސ}�� ����{�̉����D���C��w�o�ʼn�C�����D379 pp.
Koeda, K, H. Senou, C.-W. Chang and H.-C. Ho. 2019. Redescription of Liopropoma aragai (Teleostei: Serranidae), with two new confirmed records of species of Liopropoma from Taiwan. Zootaxa, 4702: 60-72.
�{���_�V�D2025�D���{�Y���ޑS��ژ^�D����܂łɋL�^���ꂽ���{�Y���ޑS��̌��݂̕W���a���Ɗw���DOnline ver. 34. https://www.museum.kagoshima-u.ac.jp/staff/motomura/jaf.html (�Q��2025-11-29)
���V�O���E�����`�G�D2015�D���{�Y���ޑS��̊w�� �ꌹ�Ɖ���D���C��w�o�ŕ��C�`��D372 pp�D
���������E�{���_�V�D2022�D�n�^��Serranidae�Ƃ���Ă������{�Y�e��̋A���C����э������ތQ�ɓK�p����W���a���̌����DIchthy, Natural History of Fishes of Japan, 19: 26-43.
Randall, J. E. and L. Taylor. 1988. Review of Indo-Pacific fishes of the Serranidae genus Liopropoma, with descriptions of seven new species. Indo-Pacific Fishes, 16: 1-47+�W pls.
���ؑ�n�D2025�D�n�i�X�Y�L�ȁi�V�́j�D�ؑ����u�E���ؑ�n�i�ҁj�Cpp. 289-290�D�������̋����� �O�d���̋��ސ}�ӁD�ɐ��D
���\ �G�D2013�D�n�^�ȁD���V�O���i�ҁj�Cpp. 757-802, 1960-1971�D���{�Y���ތ��� �S��̓��� ��3�ŁD���C��w�o�ʼn�C�`��D
�k �F��E�����L���D2025�D���m�����瓾��ꂽ�l�����L�^�̃n�^�ȋ��ރN���I�r�X�Y�L�DIchthy, Natural History of Fishes of Japan, 54: 64-68.
�c���Ε�D1922�D���{�Y���ސ}�� ��32���Cpp. 583-606 + pls. 145-150�D�ۑP�C�����D
������w�D2025�D������w�������������فC���ޕW�{�f�[�^�x�[�X �ꗗ�Dhttps://umdb.um.u-tokyo.ac.jp/DDoubutu/Fish/index.php (�Q��2025-11-30�j
�g��Y��E���\ �G�D2008�D�R��n���f�B�}��13 ���{�̊C�����D�R�Ɵ�J�ЁC�����D543 pp�D
�g��Y��E���\ �G�D2023�D�R��n���f�B�}��13 ���{�̊C�����C�����Łi2�Łj�D�R�Ɵ�J�ЁC�����D543 pp�D
van der Laan, R., R. Fricke and W. N. Eschmeyer (eds.) 2025. Eschmeyer�fs catalog of fishes: classfication. http://www.calacademy.org/scientists/catalog-of-fishes-classification/ Accessed 29 Nov. 2025.
�ʐ^�́FBSKU 138634�C138.4 mm SL�C2025�N5��19���C���m���h�юs���̓��i32.7383��N�C132.5096��E�j�C���[70���C�ނ�C�̏W�F�F�،d��E���i ���E�ˎ� �ˁi���������������j�D
�i���a�܂Ȃ݁j
2025�N10���̋�
![]()
�z�V�z�E�l���G�\Polyipnus matsubarai Schultz, 1961�i���j�g�J�Q�M�X�ڃ��l�G�\�ȁj
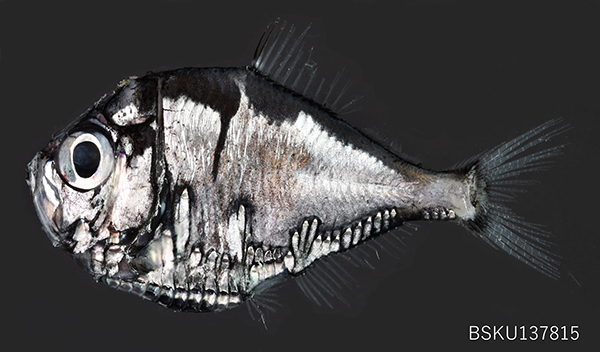
�@���l�G�\�ȃ��l�G�\���ȃz�E�l���G�\���iPolyipnus Gunther, 1887�j�͔M�т��牷�ѐ���ɕ��z���C���ɓ��{��t�B���s�����̐������m����C�T���S�C��^�X�}���C�ɂ����đ����݂��鋛�ł��iHarold et al., 2015�j�D���̑��͔w�h����N���̑O����1�̕��}���������C����������iAB�j��10�C�����㕔������iSAB�j��3�C�̑�������iL�j��1�C���h�㔭����iSP�j��1�C�\�h����������Ȃ����Ƃ�����Ƃ��Ă��܂��iSchultz, 1938; Weitzman, 1974�j�D���݂ł͐��E����34�L���킪�m���C���{�ł͂��̂����J�^�z�E�l���G�\ Polyipnus stereope Jordan and Starks, 1904�C�c�m�z�E�l���G�\ P. fraseri Fowler, 1934�C�z�V�z�E�l���G�\ P. matsubarai Schultz, 1961�C�m�R�o�z�E�l���G�\ P. spinifer Borodulina, 1979�C�}���z�E�l���G�\ P. parini Borodulin, 1979�C�X���K�z�E�l���G�\ P. surugaensis Aizawa, 1990�C�`�����v�V�z�E�l���G�\ P. ovatus Harold, 1994��7��Ɗw��������̃~�c���r�z�E�l���G�\�iPolyipnus sp.�j������Ă��܂��iHarold, 1994�G���V�E�y�����C2013�G�g�c�E�{���C2014�GHarold et al., 2016�GVourey et al., 2017�j�D
�@�z�V�z�E�l���G�\�́C���А���22-24�C�w�h�O���F�т����ɍׂ����C�\�h�㔭����iSAN�j��2�Ԗڂ�3�Ԗڂ���╠���Ɉʒu����C�㑤�������ƑO�ҊW�������Z���C�������ӂɋ������ȂȂǂ̓����ɂ�蓯�����킩���ʂ���܂��iHarold, 1994�j�D �{��͍��m���̌�������`�ł����g������Ă���C���������ɂƂ��Ă͔�r�I�g�߂ȃz�E�l���G�\�����ނł��D�{��̃z���^�C�v�ƃp���^�C�v�͌F��傩��̏W����C���{�ł͌F���̂ق��ɁC�x�͘p�C���͘p�C�y���p�ȂǂŌ����܂��D
�@�����́uPolyipnus�v�͓��{��Łu�����̔�����̂���v�Ƃ����Ӗ��ŁC���̖��̒ʂ蕠������������������ɂ����Ĕ�����̗�������܂��i���V�E�����C2015�j�D�܂��C�포���́umatsubarai�v�́C�����̋��s��w�_�w�����ߐ��Y�������ɍݐE���Ă���������㏼���m�Ɍ������ꂽ���̂ł��i���V�E�����C2015�j�D�{���̔�����͑��̃��j�g�J�Q�M�X�ڋ��ނƔ�r���đ傫���C�悭���B���Ă��܂��D�N�x���悢��Ԃł͔����킪���F�����Ă���C�����Y��ł��D �����̒��[�w�i���[ 200 m����1,000m�j�ɂ��݁C���̔��B������������p���āu�J�E���^�[�C���~�l�[�V�����v�Ƃ������@�ʼne�������āC�������猩�グ��ߐH�҂���B��Đ������Ă��܂��D���[�w�ɂ͓����ɕߐH�҂��l����F���ł�����x�̎ア������������ł���C���̌������ɂ�����Ɖe���ł��āC�ߐH�҂Ɍ������Ă��܂����Ƃ�����܂��D���̂��߁C�����̔���������点�邱�Ƃʼne�������Ďp���B���Ă��܂��D �{���̑̐F�͑��ʂ���F�C�w�ʂ����F�Ɛ������[�тł��钆�[�w�ɂ����Đ������т邽�߂Ɋl�������`�����ڗ����܂��D���̑̐F�͋��̂��ォ�猩���Ƃ��ɂ͊C������̈Èłɓ������C�������猩���ꍇ�͔�����̌��ƍ��킹�ĊC�ʑ�����̌��Ɠ������Ďp���B�����Ƃ��ł��܂��D�܂��C���̉Ȃ͉p�����uhatchetfishes�v�C���{��Łu�蕀�̂悤�ȋ��v�Ƃ����Ӗ��ɂȂ�C���̔��ɑ��G�����O�`�Ƌ�F�̑̐F���悭�\�������O�ƂȂ��Ă��܂��D �z�E�l���G�\�����ނ̈ꕔ�̎�́C�����ɂ͐[�C�ɁC��ԂɂȂ�Ɛ�C�ւƈړ�������������ړ����s�����Ƃ��m���Ă��܂��iHarold, 1994�j�D���̍s���͖{�����ނ��������钆�[�w�����\�w�̕����a�ł��铮���v�����N�g�����L�x�Ȃ��߁C�ߐH���̒Ⴂ��Ԃɕ\�w�Ɍ��������S�ɉa��H�ׂ邽�߂��Ƃ���Ă��܂��D
�@���m���̌�����ł��z�E�l���G�\�����ނ��̏W����Ă��܂����C�������{����̂Ȃ����m���V�킪������̑��J��ԋ��Ŕ������ꂽ���ɂ́C�X���K�z�E�l���G�\�̂悤�ɁC�n���̃~�}�Z�Ɉ��W���a����w����t����̂��悢��������܂���ˁD
�Q�l����
���V���G�E�y���� ���D2013�D���l�G�\�� Sternoptychidae�D���V�O���i�ҁj�Cpp. 374-380, 1836-1837�D���{�Y���ތ��� �S��̓��� ��O�ŁD���C��w�o�ʼn�C�`��D
Harold, A. S. 1994. A taxonomic revision of the sternoptychid genus Polyipnus (Teleostei: Stomiiformes) with an analysis of phylogenetic relationships. Bulletin of Marine Science, 54 (2): 428-524.
Harold, A. S., I. M. Kemp and S. K. Shore. 2016. A new species of Polyipnus (Teleostei: Stomiiformes) from the western Pacific, with comments on the P. triphanos species complex. Zootaxa, 4111 (5): 555-564.
���V�O���E�����`�G�D2015�D���{�Y���ޑS��̊w�� �ꌹ�Ɖ���D���C��w�o�ŕ��C�`��Dxv + 372 pp.
Schultz, L. P. 1938. Review of the fishes of the genera Polyipnus and Argyropelecus (Family Sternoptychidae), with descriptions of three new species. Proceedings of the United States National Museum, 86: 135-155.
Schultz, L. P. 1961. Revision of the marine silver hatchetfishes (family Sternoptychidae). Proceedings of the United States National Museum, 112 (3449): 587-649.
Vourey, E., C. Dupoux and A. S. Harold. 2017. A new species of Polyipnus (Stomiiformes: Sternoptychidae) from the Western South Pacific. Zootaxa, 4263 (3): 567-577.
Weitzman, S. H. 1974. Osteology and evolutionary relationships of the Sternoptychidae, with a new classification of stomiatoid families. Bulletin of the American Museum of Natural History, 53 (3): 327-478.
�g�c���O�E�{���_�V�D2014�D�`�����v�V�z�E�l���G�\�i�V�́j�D�{���_�V�E���Y�[��i�ҁj�Cpp. 52-53�D�����Q���œ�[�̓� �^�_���̋��ށD��������w�������������فC�������E�����Ȋw�����فC���D
�ʐ^�W�{�FBSKU 137815�C62.5 mm SL, 2024�N12��11���C���m�����m�s��������`�C�i�ہC���J��ԁC�{�艫�C���[ 250-330 m�C�̏W�ҁF����s��C����D���Y�C�V���\��C�F�،d��D
�i�͈�A��j
2025�N9���̋�

�����J�T�S Helicolenus hilgendorfii (Doderlein, 1884)�i�X�Y�L�ڃ��o���ȁj
�@����͕M�҂���D���ȃJ�T�S�̒��Ԃ̏Љ�ł��D
�@�����J�T�S�� Helicolenus Goode and Bean, 1896�͋��h�㔼���̌㉏���p�����邩�B�`�C���h�㉏���p������C���h�����ɔ�ق����邱�Ƃœ��{�Y�̃��o���ȑ����Ǝ��ʂ���܂��i���V�E�b��C2013�j�D�{���͎O��m�ɐ������C���E����9��C���{��������̓����J�T�SHelicolenus hilgendorfii (Doderlein 1884)�݂̂��m���܂��iSmith et al., 2009�G���V�E�b��C2013; Nelson et al., 2016; Fricke at al., 2025�j�D
�@�����J�T�S�͐X������F�������̑����m���݁C�ዷ�p�����B�k���݂̓��{�C���݁E���V�i�C���݁C���V�i�C�嗤�I���ӈ�C����������q���E�����哇����L�^������܂��i���V�E�b��C2013�G�{���C2025�j�D�{��͑����̏㉺�ɈÐF�����Ȃ����Ƃœ�������Ƌ�ʂ���܂��i���V�E�b��C2013�j�D���[��̍����ނ�ł悭�݂��C�悭�ނ��T�C�Y��20�`30 cm���x�ł����C�[���C��ł�60 cm�̑�^�̂��݂��܂��i�ʐ^�̂�107.9 mm SL�j�D�{��̓����Ƃ��āC���{�̈ÐԐF�̉��т�����C���h�㔼�����p������������ɔ�ق�����C�ҊW�ɍ��F�悪���邱�Ƃ��������܂��i�ΐ�C2010�G���V�E�b��C2013�G�b��E�����C2022�j�D
�@������Helicolenus�͗�����̐����Ƃ����Ӗ��ŁC�포����hilgendorfii�̓h�C�c�̊C�m�����w�҂ł���t�����c�E�q���Q���h���t�iFranz Martin Hilgendorf, 1839-1904�j�Ɉ���ŕt�����܂����i���V�E�����C2015�j�D�q���Q���h���t��1873�N����1876�N�܂Łu���ق��O���l���t�v�Ƃ��ē��{�ɑ؍݂��C�A�R�E�_�CSebastes matsubarae Hilgendorf, 1880�Ȃǖ����������{�Y���ނ͑����C���{�̋��w�E���Y�w�̕��ƌ����Ă��܂��D�i���V�E�����C2015�j
�@�{��͌��o�����������Ƃ���m�h�O���ƌĂ�܂��D�m�h�O���Ƃ����������ł���A�J���cDoederleinia berycoides (Hilgendorf, 1879) ��A�z���܂����C�֓��ł͖{��̂��Ƃ��w���悤�ł��i�ΐ�C2010�j�D�A�J���c�ɂ��Ă�2023�N11���ɏЉ�Ă���̂ŁC��������Q�Ƃ��Ă��������iLINK �j�D
�@�{��͐H�p�Ƃ��ė��ʂ��Ă���C���ɔ����������ł��D�M�҂��������߂������͎̂ϕt���ł��D�J�T�S��o�����l�C�ϕt���ɂ���Ƃق�ق�ƌ��̒��Őg������C���̖������������܂��D���m���ł��ނ邱�Ƃ��ł���̂ŁC�ނ����ۂɂ͂��АH���Ă݂Ă��������D�M�҂������ނ��Ă݂������̂ł��D
���p����
Fricke, R., Eschmeyer, W. N. & Van der Laan, R. (eds.). 2025. Catalog of fishes: genera, species,
reference. (27 August 2025)
�ΐ�ᩏ́D2010�D�ނ肪�A�����A�C�������Ɗy�����Ȃ�I�C�̋���}�ӁD�������@�C�����D400 pp.
�b��ÍW�E�����]���D2022�D���o���ȁD���V�O���i�ҁj�Cpp. 198-208�D���w�ق̐}��Z ���{���ފف`���k�Ȏʐ^�Əڂ�������`�D��6���i����j�D���w�فC�����D
�{���_�V�D2025�D���{�Y���ޑS��ژ^�D����܂łɋL�^���ꂽ���{�Y���ޑS��̌��݂̕W���a���Ɗw���DOnline ver. 32. (29 August 2025)
���V�O���E�����`�G�D2015�D���{�Y���ޑS��̊w���ꌹ�Ɖ���D���C��w�o�ŕ��C�`��Dxvi + 372 pp.
���V�O���E�b��ÍW�D���o���ȁi�V�́j�D���V�O���i�ҁj�Cpp. 668-681, 1933-1939�D���{�Y���ތ��� �S��̓��� ��O�ŁD���C��w�o�ʼn�C�`��D
Nelson, J. S., T. C. Grande and M. V. H. Wilson. 2016. Fishes of the world. 5th ed. John Wiley and Sons, Hoboken. xli + 707 pp.
Smith, P. J., C. D. Struthers, C. D. Paulin, S. M. McVeagh and R. K. Daley. 2009. Shallow genetic and morphological divergence among seaperches in the South Pacific (family Scorpaenidae; genus Helicolenus). Journal of Fish Biology, 74: 1104-1128.
�ʐ^�W�{�FBSKU 133688, 107.9 mm SL, 2023�N5��22���C���m���h�юs���m���ߊC�C���[350 m�C�ފl�C�̏W�ҁF�F�،d��C�����C�^�D
�i�吼�q�V�j
2025�N8���̋�
![]()

�A�� Plecoglossus altivelis altivelis (Temminck and Schlegel, 1846)�i�L���E���E�I�ڃA���ȁj
�@�݂Ȃ���C����ɂ��́D�y�����ނ胉�C�t�i���Ă��܂����D����͉Ă̐�ނ�Ől�C�̂��鋛�u�A���v�ɂ��ďЉ�����Ǝv���܂��D
�@�A����Plecoglossus �ɂ͝��ꂽ��Ƃ����Ӗ�������C���̖��̒ʂ�t�����ނ�H�ׂ邽�߂ɔ��B�������Ɛ�O�������Ă��܂��i���V�E�����C2015�j�D �L���E���E�I�ڂ̃A���ȃA�����iPlecoglossidae: Plecoglossus�j�ɕ��ނ����{��́C�]���T�P�� Salmonidae �̈��Ƃ���Ă��܂������CJorden (1923)�ɂ���ăA���� Plecoglossidae ������Ĉȍ~�C1��1��1��Ƃ��Ĉ�����悤�ɂȂ�܂����i���C1990�j�D�������C�A���̌n���W�ɂ��Ẳ��߂͈�肵�Ă��炸�C�L���E���E�I�� Osmeridae �Ɉʒu�Â����邱�Ƃ�����܂��i����E�|���C2006�G�����E�k��C2011�G�גJ�C2013�j�D�܂��C����܂ň��킪���Ȃ��ƍl�����Ă����A���ł����CNishida (1988) �ɂ���āC�����Ō�����A���ɂ́C�{�y�Ō�����A���Ɣ�r���āC�ȉ��̌v���`���̑��Ⴊ�F�߂��܂����F�{�y�̃A���͏c��ؐ���67-68 (vs.�����Y�ł�59-63)�C�����������ؐ���17-23 (vs. 14-16)�C��������ؐ���11-14 (vs. 9-10)�C���h��𐔂�12-15�i�ʏ�14�j�mvs. 11-13�i�ʏ�12�j�n�D�ȏ�̌`������C�����Y�̃A���͕ʈ���Ƃ���C�����E�L���E�A�� Plecoglossus altivelis ryukyuensis ���L�ڂ���܂����D ���݂ł́C�{���ɂ̓A���ƃ����E�L���E�A����2���킪�F�߂��Ă��܂��D
�@�A���̓����Ƃ��āC�w���݂͐��������I���[�u�F�ŁC�����͋┒�F�C���h�㕔�ɑȉ~��̉��F���������C�e�h�͒W�����F�����Ă��܂��D �����āC�A���͒W�����ł͂���܂����C�ꐶ��W���Ő�������̂ł͂Ȃ��C�W���ƊC���s��������ʂ���V���s���܂��i�n�ӁC2020�j�D �A���̈ꐶ�́C�͐�Ő��܂�C�C�֍~��ėc���ɂ܂Ő���������C�Ăщ͐�ɖ߂萬���܂Ő������ĎY�����s���܂��D���̂悤�ɒW������C�C�C����W���ւƈړ������V�𗼑���V�ƌ����܂��i���V�C2022�j�D �܂��C�A���̏ꍇ�͉͐�ŎY�����s���C�Z�����Ԃśz�����邽�߁C�W����������V�ƌĂ�܂��i�n�ӁC2020�j�D
�@�A���͓��{�ł͖k�C�������ȓ삩����B�ɂ����čL�����z���C���{���O�ł́C���N��������x�g�i���k���ł��������m�F����Ă��܂��i���V�C2022�j�D �����āC�͐�̏㗬���璆�������ȌC�_���C��Ղ��I��̐��╣����Ȑ����n�Ƃ��Ă��܂��D �A����ނ�ۂɂ͂����̏ꏊ�֍s���Ă݂�Ƃ悢�ł��傤�D �A���ނ�͑S���{�I�茠���J�Â����ȂǃX�|�[�c�t�B�b�V���O�Ƃ��Ă��l�C�������C�l�X�ȏꏊ�ōs���Ă��܂��D ���m���ł͐m�����l���\�삪�A���ނ�ł̐l�C�̃X�|�b�g�ƂȂ��Ă��܂��D �A���ނ�Ń��W���[�Ȏ�@�ƌ����ΗF�ނ�ł����C����̓A���̏K�������܂����p���Ă����@�Ȃ̂ł��D �A���͂Ȃ��ӎ��̋������Ȃ̂ŁC�ނ�j�Ɂu�F�A���v�ƌĂ�鐶�����A���������|���ĉj�����邱�ƂŁC�Ȃ��ɓ����������U�����ɂ����A����߂܂���Ƃ������̂ł��D �ŋ߂ł̓A���C���O�ƌĂ��C�F�A���̑���Ƀ��A�[�ŗF�ނ��������@���b��ƂȂ��Ă��܂��D ������̏ꍇ�͗F�A����p�ӂ���K�v���Ȃ����߁C�ʏ�̗F�ނ�ɔ�ׂĎ�y�ɒ��킷�邱�Ƃ��ł��܂��D �A���ނ�������ł����C��͂��Ԗ��͓I�Ȃ̂͂��̂��������D����ł̓A���̗������������Љ�����Ǝv���܂��D �A���̐H�ו��̒�Ԃ͉��Ƃ����Ă����Ă��C���m�����̃C�x���g�ł��悭����Ă��܂��ˁD���Ɋ܂ۂ̃L���E���̂悤�ȁC�X�C�J�̂悤�ȑu�₩�ȍ���Ɖ��̒��悢������ς��C�Ă����������Ă�����i�ƂȂ��Ă��܂��D ���ɂ��������݂��т�ØI�ρC�V�Ղ�Ȃǂ����Ɏ̂Ă������ł��ˁD �A�������͎�ނ��L�x�ŁC�ǂ���ƂĂ������������߁C�@�����ΐF�X�ȗ�����H�ׂĂ݂Ă͂������ł��傤���D�Ă̕���������Ă݂Ă��������D
�Q�l����
�גJ�a�C�D2013�D�A���ȁD���V�O���i�ҁj�Cpp. 360, 1832�D���{�Y���ތ����D�S��̓���D��O�ŁD���C��w�o�ʼn�C�`��D
����b��N�E�����O�F�D2006�D�A���̌Q�̍\����͂ɂ�����i�W�Ƃ��̍����I�Ӌ`�D���Y�����E����@�\�����C�ʍ�5���F187-195.
��� �ہD1990�D�A���̕��ފw�I�ʒu�D���Y���B�C38: 199-200.
��� �ہE�F�� ���D2020�D6-2 ���C� �ʁE�K���N���E�s�ؖ����i�ҁj�Cpp. 57-62�D���ފw�D�P���Ќ����t�C�����D
Jordan, D. S. 1923. A classification of fishes: including families and genera as far as known. Stanford University Publications. Biological Sciences, 3: 79-243.
���� �����E�k�� �_�D2011�D�T�P���ڋ��ނ̐����w�I�n�����ށD�O�O��w�_�w�����Ȋw���w�p�C13: 23-28.
���V�O���D2022�D�A���ȁD���V�O���i�ҁj�Cpp. 126-127�D���w�ق̐}�ӂy���{���ފف`���k�Ȏʐ^�Əڂ�������`�D��6���i����j�D���w�فC�����D
���V�O���E�����`�G�D2015�D���{�Y���ޑS��̊w���ꌹ�Ɖ���D���C��w�o�ŕ��C�`��Dxvi + 372 pp.
Nishida, M. 1988. A new subspecies of the Ayu, Plecoglossus altivelis, (Plecoglossidae) from the Ryukyu Islands. Japanese Journal of Ichthyology, 35: 236�|242.
�n� �r�D2020�D16-4 ��V�C� �ʁE�K���N���E�s�ؖ����i�ҁj�Cpp. 213-218�D���ފw�D�P���Ќ����t�C�����D
�ʐ^�W�{�FBSKU 130563�C95.3 mm SL, 2021�N7��3���C���m���y�������s�ȕz���D
�i���o�� ���j
2025�N7���̋�

�C�g�q�L�n�i�_�C Tosanoides filamentosus Kamohara, 1953 �i�y���J�ڃn�i�_�C�ȁj
�@�C�g�q�L�n�i�_�C�� Tosanoides Kamohara, 1953 �͔w�h���𐔂�10�C�w�h��1�����Œ��C�w�h������L�����Ȃ��C�\�h��3���ő�3�����Œ��C�\�h����͐��{�L������C���h����͕��}���Ȃ��C���h���[���p��������E��������͐L������C�Ғō�����10+16�C�����͋늲������Ɉʒu���C�w�h����I���܂����Ђ͍ג������ƂȂǂ̓����̑g�ݍ��킹�ŗe�Ղɓ��ȑ����Ǝ��ʂ���܂��iKamohara, 1953�GKatayama and Masuda, 1980�G���\�C2013�GGill 2021�j�D�{���͓����������㋳���̊����������m�ɂ��C1952�N�ɍ��m�����m�����瓾��ꂽ�C�g�q�L�n�i�_�C Tosanoides filamentosus Kamohara, 1953���^�C�v��Ƃ��ĐV���Ƃ��Đݗ�����܂����D�܂��C���̑����͓y���̖��������C���m�ɉ��̂���w���ł��D
�@�{���ɂ͓��{�ɐ�������C�g�q�L�n�i�_�C T. filamentosus�C�L�V�}�n�i�_�C T. flavofasciatus Katayama and Masuda, 1980�̂ق��C�n���C��������T. obama Pyle, Greene and Kosaki, 2016�C�p���I����у~�N���l�V�A����T. annepatrice Pyle, Green, Copus and Randall, 2018�C�u���W���̃Z���g�|�[���Y���b�N����T. aphrodite Pinheiro, Rocha and Rocha, 2018�C�I�[�X�g�����A��j���[�J���h�j�A����T. benetti Allen and Walsh, 2019�����ꂼ�����C����6�L���킪�m���Ă��܂��iKamohara, 1953; Katayama and Masuda, 1980; Pyle et al. 2014, 2016; Pinheiro; 2018; Anderson, 2018, 2022; Allen and Walsh, 2019; Parenti and Randall, 2020; Tea et al., 2025�j�D�܂��C�C�g�q�L�n�i�_�C�͔w�h��1�����������Ƃ���e�Ղɓ�������Ǝ��ʂ���܂��iAllen and Walsh, 2019�j�D
�@�{���̐��Ԋw�I�����Ƃ��āC�[��̊�ʑтɒP�Ƃ܂��͌Q��Ő������邱�Ƃ��m���Ă��܂��D�Ⴆ�C�C�g�q�L�n�i�_�C�̐������͐��[40 m�Ȑ[�̊�ʑтŁC���̐��[�т���͓����̃L�V�}�n�i�_�C T. flavofasciatus��}�_���n�i�_�C Odontanthias borbonius (Valenciennes, 1828)�C�V�L�V�}�n�i�_�C Callanthias japonicus Franz, 1910�C�Q�b�R�E�X�Y���_�C Chromis tingting Tea, Gill and Senou, 2019�Ȃǂ̋����m�F����Ă��܂��i���\�C1993�G���\�E����C1999; Tea et al., 2019�j�D �C�g�q�L�n�i�_�C�͓��{�����ł͍��m���C�x�͘p�i�É����吣��j�C���͘p�i�É����M�C�E�x�ˁC�ɓ��哇�j�C���{���O����̓n���C-�V�c�C�R��̂ݒm���Ă��܂��iKamohara, 1953; Gooding, 1980; ���\�C1993�C2013�j�D���̃n���C-�V�c�C�R��̋L�^�́C�n���R�b�N�C�R�i�k��30�x15���C���o178�x50���j�ō̏W���ꂽ1�̂Ɋ�Â����ꂽ���̂ł����iGooding, 1980)�D���̌�CPyle et al. (2016)�͖{�W�{�̓���ɂ��ċ^���悵�Ă��܂��D�������CGooding (1980)�ŕ��ꂽ�W�{�͌������Ȃ����߁C�{�킪�{���Ƀn���C�����k���̓V�c�C�R����ɕ��z���邩�̔��f���s���ɂ͒lj��W�{�̍̏W���]�܂�܂��D
�@���̎ʐ^�̂�2023�N�̏t�Ɏ��ˎs����l���`�ɑD���\���鏼����Ǝ��ɂ���Ēފl����C�����Ɋ���܂����D�W�{�ɂɂ̓C�g�q�L�n�i�_�C�̃z���^�C�v����������Ă��܂����C���̌�lj��W�{�͂���܂���ł����D�܂��C���R�̂��ƂȂ��瓖���̓z���^�C�v�����`�q�ؕЂ��̎悳�Ă��Ȃ��������߁C73�N�U��Ɋ��ꂽ�{�W�{�͈�`�q�T���v���E�J���[�ʐ^���������ō��̌̂ƂȂ�܂����D�������ɂ̓C�g�q�L�n�i�_�C���͂��߁C����E�����i2024�j�ŕ����c�L�q�n�i�_�C�C�Ö�ق��i2024�j�ŕ����A�J�z�V�L�c�l�x���ȂǑ���ɓn�蒲���ւ����͂����������Ă���܂��D�����ɂ�1�N�ɐ���̂̋��ނ���������܂����C���������͂��ߑ����̕��̂����͂Ŋ��ꂽ���ނ����Ȃ�����܂���D�{�e�������܂��ĊF�l�ɉ��߂Ă���\���グ�܂��D
���p����
����E�����L���D2024�D���m���Ƙa�̎R�����瓾��ꂽ�k���L�^���܂ރc�L�q�n�i�_�C�DIchthy, Natural History of Fishes of Japan, 40: 17-21.
Allen, G. R. and F. Walsh. 2019. Tosanoides bennetti, a new species of anthiadine fish (Pisces: Serranidae) from the Coral Sea. aqua, International Journal of Ichthyology, 25: 25-34
Anderson, W. D., Jr. 2018. Annotated checklist of anthiadine fishes (Percoidei: Serranidae). Zootaxa, 4475: 1-62.
Anderson, W. D., Jr. 2022. Additions and emendations to the annotated checklist of anthiadine fishes (Percoidei: Serranidae). Zootaxa, 5195: 567-578.
Gill, A. C., J. J. Pogonoski, G. I. Moore and J. W. Johnson. 2021. Review of Australian species of Plectranthias Bleeker and Selenanthias Tanaka (Teleostei: Serranidae: Anthiadinae), with descriptions of four new species. Zootaxa, 4918: 1-116.
Gooding, R. M. 1980. Exploratory fishing on the Hancock seamounts by the Townsend Cromwell, 1976-79. Pages 241-250 in R. W. Grigg and R. T. Pfund, eds. Proceedings of the Symposium on Status of Resource Investigations in the Northwestern Hawaiian Islands. University of Hawai�ei Sea Grant College Program, Honolulu.
Kamohara, T. 1953. Marine fishes newly found in Prov. Tosa, Japan, with descriptions of a new genus and species. Research Reports of Kochi University, 2 (11): 1-10, 2 pls.
Katayama, M. and H. Masuda. 1980. Tosanoides flavofasciatus, a new anthiine fish from Sagami Bay, Japan, and the Tonga Ridge, Melanesia. Japanese Journal of Ichthyology, 27(1): 51-55.
Parenti, P. and J. E. Randall. 2020. An annotated checklist of the fishes of the family Serranidae of the world with description of two new related families of fishes. FishTaxa, 15 (1): 1-170.
Pinheiro, H. T., C. R. Rocha and L. A. Rocha. 2018. Tosanoides aphrodite, a new species from mesophotic coral ecosystems of St. Paul�fs Rocks, Mid Atlantic Ridge (Perciformes, Serranidae, Anthiadinae). ZooKeys, (786): 105-115.
Pyle, R. L., B. D. Greene and R. K. Kosaki. 2016. Tosanoides obama, a new basslet (Perciformes, Percoidei, Serranidae) from deep coral reefs in the northwestern Hawaiian Islands. ZooKeys, (641): 165-181.
Pyle, R. L., B. D. Greene, J. M. Copus and J. E. Randall. 2018. Tosanoides annepatrice, a new basslet from deep coral reefs in Micronesia (Perciformes, Percoidei, Serranidae). ZooKeys, (786): 139-153.
���\ �G�D1993�D�����̋� �C�g�q�L�n�i�_�C�D�ɓ��C�m�����ʐM�C4 (2): 1
���\ �G�D2013�D�n�^�ȁD���V�O��(��)�Cpp. 757-802, 1960-1971. ���{�Y���ތ��� �S��̓��� ��O��.���C��w�o�ʼn�C�`��.
���\ �G�E���� �C�D1999�D�����̋� �o���n�i�_�C�i�c���j�D�ɓ��C�m�����ʐM�C10 (12): 1
Tea, Y.-K., A. C. Gill and H. Senou. 2019. Chromis tingting, a new species of damselfish from mesophotic coral ecosystems of southern Japan, with notes on C. mirationis Tanaka (Teleostei: Pomacentridae). Zootaxa, 4586: 249-260.
Tea, Y.-K., T.-L. Sih, F. Walsh, T. Bennett, L. A. Rocha, G. Bardout, A. Hay, K. Parkinson, S. Reader, I. J. Riley, J. J. Pogonoski, J. W. Johnson, G. I. Moore, A. C. Gill, G. F. Galbraith, B. J. Cresswell, A. S. Hoey, B. W. Frable, B. P. Brooke and R. J. Beaman. 2025. New records of fishes from the Coral Sea Marine Park, Australia. Coral Reefs:1-48. [First appeared online, pp. 1-48, on 16 June 2025.]
�Ö�`��E�F�،d��E����E�����L���D2024�D���˖������瓾��ꂽ���m�����L�^�̃A�J�z�V�L�c�l�x���DIchthy, Natural History of Fishes of Japan, 44: 26-30.
�ʐ^�W�{�FBSKU 133733, 81.6 mm SL, 2023�N5��25���C���m�����ˎs����l���C���[120 m�C�ފl�F������ƁC�W�h�E�B�e�F����D
�i����j
2025�N5���̋�
![]()

�c�}���}�c�J�TMyripristis greenfieldi Randall and Yamakawa, 1996�i�L�����_�C�ڃC�b�g�E�_�C�ȁj
�@����͍��m���Ń_�C�r���O�|�C���g�Ƃ��ėL���Ȕ����ɂ䂩��̂���Ԃ����̏Љ�ł��D
�@�A�J�}�c�J�T��Myripristis�̓C�b�g�E�_�C�ȃA�J�}�c�J�T���ȁiHolocentridae; Myripristinae�j���\������1���ł���C�t�����X�̔����w��Georges Cuvier �ɂ��Myripristis jacobus Cuvier, 1829���^�C�v��Ƃ��Đݗ�����܂����iRandall and Yamakawa, 1996�j�D�{���́C�O��m�̔M�сE���M�ъC��𒆐S�ɐ������C72���`��30�L���킪�m���Ă��܂��iRandall and Greenfield, 1996; Nelson et al., 2016; Fricke at al., 2025�j�D���̂����C���{�ł�15��C���m���ł�7�킪�L�^����Ă��܂��i���c�ق��C1996�G���V�ق��C2001�G�сC2013�G�{���C2025�j�D
�c�}���}�c�J�TMyripristis greenfieldi Randall and Yamakawa, 1996�́C�A�J�}�c�J�T���ȃA�J�}�c�J�T���ɑ����C�X��ʐ����ނ̕��ނŒ�����John E. Randall���m�Ɩ{������OB�̎R��@�����ɂ��C�z���^�C�v�ƃp���^�C�v�̍��v12�W�{�ɂ��ƂÂ��V��L�ڂ���܂����D���̂����C�{�������ɂ̓p���^�C�v��5�W�{����������Ă��܂��i�ʐ^�j�D���L�ڂł́C���m�������S�匎�������Ɨ����i�����哇�C����j����L�^����C�z���^�C�v�ƃp���^�C�v5�W�{�͔������ӂł̎h���ԋ��ō̏W����܂����D���L�ڈȌ���C���h�i�P�����B�j�Ɠ��{����lj��L�^������C���{�����ł͏��}�������C���͘p�ȓ�̓���{�̑����m�݂��甪�d�R�����i�Ί_���j�܂ōL�����z���܂��iRandall and Yamakawa, 1996�G�сC2013�G�]���E�{���C2016; Nair and Dineshkumar, 2016�G�{���C2025�j�D
�@�{��͋��h���������C�����L�E�ؐ���28-29(�ʏ�29)�C���������̑̑��؉��ӂ̏�����35-52�C���{�̎���1�C�O�����O�[���킸���ɉ���C�ҊW���Ƌ��h�������ӂ����F�C�w�h���̕h����[�t�߂��ԐF�C���h�O�������F �Ƃ������������瓯������Ǝ��ʂł��܂��iRandall and Greenfield, 1996; Randall and Yamakawa, 1996�j�D�{��͓���Myripristis randalli Greenfield, 1974�Ɨގ����܂����C�������w�h�Œ�����2.0-2.25�{�ivs. 1.8-2.0�j�C�������\�h��3����1.85-2.1�{�ivs. 1.5-1.8�j�C�����đ��������̑̑��؉��ӂ̏�����35-52�ivs. 21-30�j�ł��邱�Ƃ��疾�ĂɎ��ʂ���܂��iRandall and Greenfield, 1996�j�D �@
�@�ʐ^���̕W�{�́C�����ɂ����ĕM�҂���ނ�Œފl�������̂ŁC���n�̕��ʎ�ł���i�~�}�c�J�TMyripristis kochiensis Randall and Yamakawa 1996�ɍ�����1�̂��̏W���܂����D�A�J�}�c�J�T�����ނɂ͊O�������ɗގ�����킪�����܂܂�C�����̎��ʂ�������ߌ듯�肳�ꂪ���ȕ��ތQ�ł�����܂��D���ɁC�A�J�}�c�J�T�Ɠ��肳�ꂽ�W�{�ɂ͕����킪��������ꍇ�������悤�ł��D������ނ�Ȃǂō̏W����@�����܂����玝���A���Ď퓯��ɒ��킵�Ă݂Ă��������D���͒������킩������܂���D
���p����
�]���c��E�{���_�V�D2016�D�����ɂ�����C�b�g�E�_�C�ȋ��ޑ��DNature of Kagoshima, 42: 57-112.
Fricke, R., W. N. Eschmeyer and R. van der Laan (eds.). 2025. Eschmeyer�fs catalog of fishes: genera, species, references. (http://researcharchive.calacademy.org/research/ichthyology/ catalog/fishcatmain.asp) (7 May 2025)
�с@���`�D2013�D�C�b�g�E�_�C�ȁCpp. 579-591, 1897-1899�D���V�O���i�ҁj���{�Y���ތ����@�S��̓���D�� 3 �ŁD���C��w�o�ʼn�C�`��D
���c�q�@�E�R��@���E��c���v�E�^��O�Y�E�����@�j�E�吼�M�O�D1996�D���m�������̋��ޑ��@�s���Ɛ��ԂɊւ���L�q�𒆐S�Ƃ��āD���m��w�C�m�������猤���Z���^�[�����C(16): 1-177�D
�{���_�V�D2025�D���{�Y���ޑS��ژ^�D����܂łɋL�^���ꂽ���{�Y���ޑS��̌��݂̕W���a���Ɗw���DOnline ver. 30. (28 Apr. 2025)
Nair, R. J. and S. Dineshkumar. 2016. New distributional records of three soldier fishes (Pisces: Holocentridae: Myripristis) from Indian waters. Marine Biodiversity Records, 9: 1-5.
���V�O���E���c�g�F�E�R���k��E���c�����i�ҁj�D2001�D�ȕz�������̋� �W���x�G�U������}���{�E�܂ŁD�C�V�فC���D300 pp.
Nelson, J. S., T. C. Grande and M. V. H. Wilson. 2016. Fishes of the world. 5th ed. John Wiley and Sons, Hoboken. xli + 707 pp.
Randall, J. E. and D. W. Greenfield. 1996. Revision of the Indo-Pacific holocentrid fishes of the genus Myripristis, with description of three new species. Indo-Pacific Fishes, (25): 1�|61.
�ʐ^�F�{�����������̃c�}���}�c�J�TM. greenfieldi�̃p���^�C�v�W�{�iBSKU 82130-82134�j LINK
�W�{�ʐ^�FBSKU 137109�C91.0 mm SL�C���m�������S�匎�������C2024�N9��26-27���C�̏W�E�ʐ^�B�e�F���i ���ق��D
�i���i ���j
2025�N3���̋�


���j�O�`�c�m�U�� Trigonognathus kabeyai Mochizuki and Ohe, 1990�i�c�m�U���ڃJ���X�U���ȁj
�@���j�O�`�c�m�U���͏��^�̐[�C���T���ނ̋H��ŁC1986�N�ɘa�̎R���������i���[330 m�C�z���^�C�v�j�Ɠ��������a�����i���[360 m�C�p���^�C�v�j�����g�ԋ��œ���ꂽ2�W�{����ɁC1990�N�ɐV���V��Ƃ��ċL�ڂ���܂����iMochizuki and Ohe, 1990�j�DShirai and Okamura (1992)��1988�N�ɍ��m�s��������`�̑��J��ԋ��œ���ꂽ�{��̂P�W�{�iBSKU 44653, �S��258 mm�C���m���ł̍ŏ��̕W�{�j�̍��i�n�Ƌؓ��n���r��U���C�{�̋@�\�`�Ԃ⌻�݂̃J���X�U���Ȃ̑����Ƌ߉��ł��邱�Ƃ𐄑����C�����͋��ɃJ���X�U�����Ȃɕ��ނ���܂����D�{��͌����傫���C�������{�ɂ͉s���C���Ȃ������ג��������܂�ɕ��т܂��D���{�͎O�p�`�ōׂ����C��{�͓������痣��đO���֓ˏo���邽�߁C�{���o��Ɩ{�Ȃ̒��ł��Ɠ��̕��e�ɂȂ�C�����~�c�N���U���Ɏ��Ă��܂��D���j�O�`�c�m�U�����~�c�N���U���Ɠ��l�ɁC�f�����{��ˏo���Ċl�����s�����ő�����ߐH�l���iclutching-type feeder�j�ƍl�����܂����D����C���̃c�m�U���Ȃ̎�́C���듪�̏�{���Ɖs������̉��{�����g���āC����P��Ȃ���a�̓����̓������{���Ŋ��ݎ�邽�ߕߐH�l�����قȂ�܂��D���̌�CYano et al. (2003) �͌F���ō̏W���ꂽ�{���36�W�{�Ə��}�������ߊC�Ń��o�`��V�}�K�c�I�ނɕߐH���ꂽ3�W�{�C�^�C�v��2�W�{����ɁC�{��̐�����n�C�H�������܂����D�{��̃I�X�͑S��43 cm�C���X�͑S��52 cm�Ő��n���C�ő�őS��54 cm�ɒB���܂��D�{��͂����Ƀn�_�J�C���V�ނ�H�ׂĂ���C�ߐH�̂̑S��30~40%�̑̃T�C�Y�̉a���ۓۂ݂ɂ��Ă��܂����D�������C�ߐH���ꂽ���ނɂ͖{��̉s�����̐Ղ��t�����̂�����C�S�����̂Ȃ����̂������CYano et al. (2003) ��Shirai and Okamura (1992)�����������ߐH�l���Ƃ͈قȂ��āC�j���Ȃ���傫�������J���ĉa���l�銷���^�̕ߐH�l���iram-feeder�j�ł͂Ȃ����ƍl���܂����D������������C�{���2�ʂ�̗l�����g�������Ă���̂�������܂���D
�@���j�O�`�c�m�U���͎�v�ȉa�ł���n�_�J�C���V�ނƓ��l�ɁC�\�w���璆�[�w�i���[200 m����1,000 m�j�œ��������ړ����s���ƍl�����Ă��܂��iYano et al., 2003; Claes et al., 2014�j�D�{��͍̑̂��F�ŁC�J���X�U���Ȃ̑���Ɠ��l�ɁC���Z�����F�̕����ɂ͑����̔����Ȕ����킪���z���C���[�w�ł̃J�E���^�[�C���~�l�[�V�����̖����┭�����ʂ̕��z�̑��l������C��⎓�Y�Ԃ̃V�O�i���C�G����̖h��i�w�h���̎��́j�ȂǁC���̖�������������Ă��܂��iDuchatelet et al., 2021�G���C2022�j�D����ނŔ��������́C�c�m�U���ڂ̃����C�U���ȁC�J���X�U���ȁC�����ăI���f���U���Ȃ�3�ȂɌ����܂��i���C2022�j�D�����̃T���ނ̔����͂��ׂĐF�ŁC�J���X�U���ȃJ���X�U�����ł́C�Z�����e���W������Ƃ������V�t�F�����|���V�t�F���[�[�����Ɣ������܂����iMizuno et al., 2021�G���C2022�j�D�G�r�ނ�J�C�A�V�ނ����Z�����e���W���́C�H���A����ʂ��ăJ���X�U�����̂悤�ȕߐH�҂ɂ��C�J�C�A�V�ނ��a�Ƃ���n�_�J�C���V�ނȂǂ���ԐړI�Ɏ�荞�܂��悤�ł��D���̂��߁C��������T���ނ́u�����������^�C�v�v�Ɨ\�z����Ă��܂��i���C2022�j�D
�@���[�w�́u�g���C���C�g�]�[���v�ƌĂ�C�����͕\�w����̂킸���Ȍ����͂����ł��D�J���X�U���Ȃ���C�U���Ȃ̔�������T���ނ��C���̓���Ȋ��ւ悭�K��������������܂��iClaes et al., 2014�j�D�J���X�U���Ȃ͊�̏㉏�ɔ�������Ƃ���ɗאڂ��锭����������C���̕����͓����ɏ������͂����ʂ����m���āC�J�E���^�[�C���~�l�[�V�����̔�����̌��ʂ̃R���g���[���₻�̉����ɊW����ƍl�����Ă��܂��iClaes et al., 2014�j�D���j�O�`�c�m�U���ł͂��̔������悪�傫���C�J���X�U�����̎�ɔ�ׂĖڗ����܂��D �܂��C�{��̊ዅ���ʁi��������j�ɂ������e��̕��z�Ɩ��x�́C���I�Ȍ����ɂ��ޓ����������C�����ɂ͊C��t�߂ɂ��邱�Ƃ��x�����܂��iClaes et al., 2014�j�D����͓����ɖ{�킪��g�Ԃŋ��l����闝�R�ł��傤�D �{��͓��{�ł͏x�͘p����y���p�܂ł̖{�B�Ǝl���̑����m�݉��C�F�{���̓��V�i�C�C�����ď��}�������ߊC����L�^������C���O�ł͑�p�ƃn���C�������������܂��i�x�]�E�c���C2015�G�O�V�C2024�j�D�̏W���[�͕\�w���琅�[ 1,000 m�܂ŁC���[�w�𒆐S�ɏo�����܂��D
�@Shirai and Okamura (1992)����U�p�Ɏg�p����BSKU�̕W�{�́C�����C�m1�N����1988�N4���ɍ��m�s�̌���������֍̏W�ɍs�����ۂɏE�������̂ŁC�������̃R���N���[�g�̏��ɂP�̂��������Ă��܂����D���̎��_�ł͖��L�ڎ�ł������C�]�����m�i�����C������w�j���L�ڗ\��Ƃ킩��܂����D���̌�C1990�N�Ɏ����k�C����w���Y�w���̔��m�ے��i�w�����ۂɁC�������̐�y�ł��������䎠���m�ɂ��̕W�{�̏���`���āC���̌�ɔ�r��U����ĉ������搶�Ƃ̋����_���ɂȂ��Ă��܂��D���̎ʐ^�͖̂{�������̌F�،d��N���P�N�̎��Ɍ�������������̎i�ۂ֏�D���C�̏W����2�W�{�̂�����1�ł��D
���p����
Claes, J. M., J. C. Partridge, N. S. Hant, E. Garza-Gissholt, H.-C. Ho, J. Mallefet and S. P. Collin. 2014. Photon hunting in the twilight zone: visual features of mesopelagic bioluminescent sharks. PloS One, 9(8): e104213. DOI:10.1371/journal.pone.0104213 LINK
Duchatelet, L., J. M. Claes, J. Delroisse, P. Flammang and J. Mallefet. 2021. Glow on sharks: state of the art on bioluminescence research. Oceans, 2: 822-842. LINK
�x�] ��E�c�� ���D2015�D�x�͘p�̐[�C��g�Ԃō̏W���ꂽ���j�O�`�c�m�U�� Trigonognathus kabeyai �ɂ��āD���җތ������C(51): 17-18.
�O�V �ɁD2024�D�J���X�U���ȁCpp. 17-18�D���{ ���E�{���_�V�i�ҁj�D���{�̐[�C���}�ӁD�R�ƌk�J�ЁC�����D LINK �@���}�P�C�̖{ note
Mizuno, G., D. Yano, J. Paitio, H. Endo and Y. Oba. 2021. Etmopterus lantern sharks use coelenterazine as the substrate for their luciferin-luciferase bioluminescence system. Biochem. Biophys. Res. Commun., 577: 139-145. LINK
Mochizuki, K. and F. Ohe. 1990. Trigonognathus kabeyai, a new genus and species of the squalid sharks from Japan. Japan. J. Ichthyol., 36(4): 385-390.
���T��D2022�D���E�̔��������D���ށE���ԁE�������J�j�Y���D���É���w�o�ʼn�C���É��D433 pp. LINK
Shirai, S. and O. Okamura. 1992. Anatomy of Trigonognathus kabeyai, with comments on feeding mechanism and phylogenetic relationships (Elasmobranchii, Squalidae). Japan. J. Ichthyol., 39(2): 139-150.
�a�c�p�q�E�O�ؗ����E�����E�{���_�V�D2019�D�F�{���V���s�ߊC���瓾��ꂽ�F�{�����L�^���܂ދ��ށD�F�{�쐶����������C(9): 17-24.
Yano, K., K. Mochizuki, O. Tabeta and K. Suzuki. 2003. Further description and notes of natural history of the viper dogfish, Trigonognathus kabeyai from the Kumano-nada Sea and the Ogasawara Islands, Japan (Chondrichthyes: Etmopteridae). Ichthyol. Res., 50: 251-258.
�ʐ^�W�{�FBSKU 130940�C�S�� 410 mm, �I�X�C���m�s����������C�i�ہC���J��ԁC���[ 380 m�C2021�N10��21���C�̏W�ҁF�F�،d��D
��BSKU 44653, �S��258 mm�C���m�s����������C���J��ԁC���É��C���[ 200-230 m�C1988�N4��20���C�̏W�ҁF�����L���D
�i�����L���j
2025�N1���̋�

�I�j�A�I���G�\ Bathysauroides gigas (Kamohara, 1952)�i�q���ڃI�j�A�I���G�\�ȁj
�@�I�j�A�I���G�\��1950�N4��15���ɍ��m�s��������s��i���J��ԋ��j�ō̏W���ꂽ�P�W�{�iBSKU 3852, �z���^�C�v�C242 mm SL�j�Ɋ�Â��C�A�I���G�\�� Bathysaurops Fowler, 1938�̐V��Ƃ��Ċ����������m�i1901-1972�j�ɂ��L�ڂ���܂����iKamohara, 1952�j�D�������C���̗��N�Ɋ������m�͂��̑��� Bathysauropsis Regan, 1911�̐V�Q�ٖ��ƌ��Ȃ��Ă��܂��iKamohara, 1953�j�D���̌�CBaldwin and Johnson (1996)�͖{����^�C�v��Ƃ��ĐV���� Bathysauroides ��ݗ����C�{����`���E�`���n�_�J�� Ipnopoidae �Ɋ܂߂܂����D����ɁCSato and Nakabo (2002)�� Bathysauroides Baldwin and Johnson, 1996���^�C�v���Ƃ��āC�V�� Bathysauroididae �I�j�A�I���G�\�Ȃ�ݗ����Ă��܂��D��P���҂̍����F�N���m�́C�{�������Ŋw���ƏC�m�ے����߂����āC���s��w��w�@�̔��m�ے��܂ŃA�I���G�\�Ȃ𒆐S�Ƀq���ړ��̌n���މ��W���������C�V�ȐV���̃i�K�A�I���G�\�ȃi�K�A�I���G�\���iParaulopidae, Palaurops�j��ݗ����܂����iSato and Nakabo�A2002)�D�I�j�A�I���G�\�Ȃ̓q���ڂ̃{�E�G���M�����ڂɕ��ނ��ꂽ���Ƃ�����܂������C���݂͎b��I�Ƀ~�Y�E�I���ڂɊ܂߂��Ă��܂��i���V�E�b��C2013; Near and Thacker, 2024�j�D
�@�I�j�A�I���G�\�͋H��ŁCYamakawa (1981)�ɂ�茴�L�ڈȗ��ƂȂ�lj��̂P�W�{�iBSKU 50035, 195.7 mm SL�C1980�N12��11���C���m�s��������s��j�����N���̎ʐ^�Ƌ��ɕ���܂����D���̗��N��1981�N3��23���ɂ��C��������s���3�ԖڂƂȂ�P�W�{���̏W����C�������ɖ{��̓���Ȋ�̓��ʂ�Ԗ��̐��̂Ɋւ���V�m��������܂����iSomiya et al., 1996�j�D�{��̎e�t���ɂ��Ă̒m�������Ȃ��C���{�Y�t���}�ӑ�2�łɂ� 7.5 mm SL�i�[�����̍�������j��10.1 mm SL�i�I�[�X�g�����A�ߊC�Y�j�̎e���̐��悪�f�ڂ���Ă��܂��D�ߔN�C��Ԃɕ\�w��ɕ��V���鋛�ނ̎e�t���△�Ғœ����̗c���𐅒��B�e���� "Blackwater diving" ������ɍs���Ă��܂��i�Ⴆ�CMilisen, 2020; Nonaka et al., 2021�j�D���̂悤�Ȑ����ł́C����܂őS���m���Ă��Ȃ������[�C���̎e�t���̎ʐ^�⓮��C�W�{���̏W����邱�Ƃ�����C�̐F���܂߂��e�t���̊��S�Ȍ`�Ԃ�s�������炩�ɂȂ��Ă��܂��D���N7���ɁC�����ʐ^�Ƃ̕��������v�ē��ŎB�e�����I�j�A�I���G�\�e���̓��悪youTube�Ō��J����Ă��܂����D���̌̂͂���t���̋��h�̏㔼�����N�₩�ȃI�����W�F�����Ă���C���h��O���ɍL���ăo���U�C��������ԂŁC�����Œ��������p��������Ă��܂��D�܂��C���͔̌̂w�h���\�h�C���h�͂܂��o�����Ă��Ȃ��悤�ł��D���R�i2013�j�ɂ��C�{��̕h�͔��h�C���h�C�\�h�ƕ��h�C�����Ĕw�h�̏��Ԃɔ�������ƋL�ڂ��Ă��܂��D�������琄������ƁC���̌̂̑̃T�C�Y���̒� 10 mm ���x��������܂���D
�@���̎ʐ^�W�{�͏�Ԃ��悭�C����W�{���̏��Ȃ���̑����I�j�A�I���G�\�̌����ɖ𗧂ł��傤�D
���p����
Kamohara, T. 1952. Revised description of the offshore bottom-fishes of Prov. Tosa, Shikoku, Japan. Reps Kochi Univ., Nat. Sci., (3): 1-122.
Kamohara, T. 1953. A review of the fishes of the family Chlorophthalmidae found in the waters of Japan. Japan. J. Ichthyol., 3(1): 1-6.
Milisen, J. 2020. A field guide to blackwater diving in Hawai'i. Mutual Publishing, Honolulu. 282 pp.
���V�O���E�b��ÍW�D2013�D�I�j�A�I���G�\�ȁCpp. 426, 1851-1852. ���V�O���i�ҁj ���{�Y���ތ��� �S��̓���D��3�ŁD���C��w�o�ʼn�C�`��D
Near, T.J. and C. e. Thacker. 2024. Phylogenetic classification of living and fossil ray-finned fishes (Actinopterygii). Bull. Peabody Mus. Nat. Hist., 65(1): 3-302.
Nonaka, I., J. W. Milisen, B. C. Mundy and G. D. Johnson. 2021. Blacwater diving: an exciting window into the planktonic area and its potential to enhance the quality of larval fish collections. Ichthyol.Herpetol., 109(1): 138-156. Link
���R�@�Y�D2013�D�I�j�A�I���G�\�ȁCpp. 283-284. ���R�@�Y�i�ҁj���{�Y�t���}�ӁC��2�ŁD���C��w�o�ʼn�C�`��D
Sato, T. and T. Nakabo. 2002. Paraulopidae and Paraulopus, a new family and genus of aulopiform fishes with revised relationships within the order. Ichthyol. Res., 49(1): 25-46.
Somiya, H., T. Yamakawa and M. Okiyama. 1996. Bathysauropsis gigas, a deep-sea aulopiform fish with a peculiar iris process and a pure-cone retina. J. Fish Biol., 49 (Suppl. A):175-181.
Yamakawa, T. 1981. Second record of Bathysauropsis gigas (Kamohara) from Kochi, Japan. Mem. Fac. Sci. Kochi Univ., Ser. D (Biol.), 2: 29-32.
�������@���D���E���̋M�d�f���I�j�A�I���G�\�̒t��-Black Water Dive�i2023/07/03 �v�ē��j�DYouTube�`�����l�� Ryo Minemizu
�@
�ʐ^�W�{�FBSKU 137940�C175.5 mm TL�C2025�N1��12���C���m�����m�s��������`�C�i�ہC
���J��ԁC�y���p�����C���[ 200-318 m�C�̏W�ҁF�F�،d��C�����C�^�C����s��D
�i�����L���j
|
Copyright (C) Laboratory
of Marine Biology, Faculty of Science, Kochi University (BSKU) |
{kind=link}