2026�N6���̋�

�h�E�N�c�~�~�Y�n�[�ߎ��� Luciogobius cf. albus Regan, 1940�i�n�[�ڃn�[�ȁj
�@�n�[�ȃ~�~�Y�n�[�� Luciogobius Gill, 1859 �̃h�E�N�c�~�~�Y�n�[�́C�����̃C�h�~�~�Y�n�[ L. pallidus �ƂƂ��ɑ̂̐F�f�������Ⴊ�މ������n���������ނŁC���A����ѕt�߂̈�˂�͐�̕��������ɐ������C�������C���茧�C����э��m������L�^������܂��iTomiyama, 1936�GRegan, 1940�G���ÁC1963�G����C1972�G�{�n�ق��C1976�G�����ق��C1984�G���m�ق��C2013�G����C2015�G����, 2018�G�����ق��C2024b�j�D�{���2006�N�ȍ~�̏W�Ⴊ�Ȃ��C�������C���茧�C����э��m���̃��b�h���X�g�ɂ����āC���ꂼ���� (EX)�C��Ŋ뜜IA�� (CR)�C����я��s�� (DD�j�Ɣ��肳��܂����i�R���C2014�G�����C2018�G���茧�C2022�j�D
�@�̂��ɁC�h�E�N�c�~�~�Y�n�[��C�h�~�~�Y�n�[�Ƃ��ċL�ڂ�����ɂ��Ă̍ŏ����̕Ƃ��ẮC��c�i1934�j��Tomiyama (1936)������܂��D�����͓����C���m���o�g�̓c���Ε䔎�m�ɂ���āC�~�~�Y�n�[ L. guttatus Gill, 1859 �����܂��ܒn���̈Ï��ɖ������C�����̈Í������ɓK������2���I�ɕό`�������̂ł���Ƃ���܂����i��c�C1934�G����C1972�G���ÁC1979�j�D��c�i1934�j�͓������卪���̓��A�ƘA�������˂���C�F�������Ⴊ�������ޏk�މ������~�~�Y�n�[��2�̓����Ƃ��܂����m���Ái1979�j�ɂ��C����1�͓̂��A���̒n����������Ԃŋd�����ꂽ�n�D���n�ł́C���A���Ɂu���i�V�E���v�i�����}�}�j������Ƃ̌����`�����������Ƃ����܂��i��c�C1934�j�D�܂��C���l�̗�Ƃ��āC��N�i�����}�}�j�a�̎R�������K�S�Í����i���E���{���j�̈�˂���~�~�Y�n�[���̏W���ꂽ���Ƃ������܂����i��c�C1934�j�DTomiyama (1936)�͖M�Y�~�~�Y�n�[�����ނ�C���h�C�����̔玿�ˋN�̗L���Ȃǂ���1��4����ɕ��ނ��܂����D���̒��ŁC���A���瓾��ꂽ�C�A���r�m�Ŕw�h���\�h�̕h�𐔂����Ȃ��C�����傫���Ⴊ�ޏk���V���h�����Ȃ�2�W�{�m���V�i2001�j�ɂ��C��c�i1934�j�Ɠ����W�{�n�ƁC�x����˂��瓾��ꂽ�C�̂������C�w�h���\�h�̕h�𐔂����Ȃ��C�Ⴊ�牺�ɖ��v���C�V���h�����Ȃ�3�W�{���~�~�Y�n�[ L. guttatus guttatus �̌̕ψقɊ܂߂C���ꂼ��ʁX�ɐ}�����܂����D
�@�����āCRegan (1940)�́CTomiyama (1936) �̑}�}����ɁC���A���瓾��ꂽ�W�{�� L. albus�C�x����˂��瓾��ꂽ�W�{�� L. pallidus �Ƃ��ĐV��L�ڂ��C�����i1955�j��Tomiyama (1936) �̑}�}�����p���CRegan (1940) ���������`�ԓI�����Ɛ������������āC���ꂼ��Ƀh�E�N�c�~�~�Y�n�[�ƃC�h�~�~�Y�n�[�̘a����^���܂����iDotu, 1957�G���ÁC1979�G�a��ق��C2019�j�D���̌�CDotu, (1957) ��Tomiyama (1936) ���p�����W�{���ώ@�������ʁCRegan (1940) ������������̎��ʌ`���͌̍��������ł��������ƁC�̌`��̐F�͍̏W��̎�����Ԃɂ���đ傫���ω��������Ƃ������ɁCL. pallidus �� L. albus �̓����ٖ��ƌ��܂����D�������CShiogaki and Dotsu (1976) �͗����Ғō����Ɠ����̑̒��䂩�环�ʂł���Ƃ��CL. pallidus �������܂����D�{�n�ق��i1976�j�����l�ɕʎ�Ƃ݂Ȃ��C���̍����ɍ��m���ł͗��킪�������瓾��ꂽ���Ƃ������Ă��܂��D�����́C����������ਉƐߖ펁�Ɖ��� �����������m���V����ł̒����ɂ����āC�h�E�N�c�~�~�Y�n�[���C�h�~�~�Y�n�[�Ɠ�������̏W���C����̌`�ԓI���ق��������ē��{���ފw��N��Ŕ��\�������Ƃɂ��܂��iਉƁE�����C1976�j�D
�@���̌�CKanagawa et al. (2011) �͐É������삩�瓾��ꂽ�W�{����ɁC���E�X�C�~�~�Y�n�[ L. fonticola ��V��L�ڂ��܂����D����Ƃ����̌Q�́C�É������{��C�O�d�����q��C����јa�̎R���Í��삩�������Ă��܂��i�����ق��C2018�C2019�G�a��ق��C2019�G�����C2022�G�r���E�����C2026�j�DKanagawa et al. (2011)�C���m�ق��i2013�j�C����ы���i2015�j�́C�h�E�N�c�~�~�Y�n�[�ƃ��E�X�C�~�~�Y�n�[�̎��ʌ`���Ƃ��āC����Ԋu�悪�c�o����i��҂ł͕��R�j�C�������̒���1/4�ȏ�i1/5�O��j�C��{���̑̒��䂪����12.9 %�C�W���� �}1.1 %�i10.5�}1.1%�j�C�������̑̒��䂪8.5 %�}0.3 %�i7.3�}0.8 %�j�C����ъႪ�[�����v���֊s���s���āi���v���֊s�����āj��5�������Ă��܂��D�������C�a��ق��i2019�j�͒��茧�Y�h�E�N�c�~�~�Y�n�[�ƐÉ����Y���E�X�C�~�~�Y�n�[�̕W�{���ώ@�������ʁC����Ԋu��̖c�o���x�͗��҂Ƃ��̂ɂ��l�X�ŁC�����C��{���C����є������̑̒��ɑ��銄���͗��҂ňꕔ�d�����C��̖��v���x�ɂ����ق��F�߂��Ȃ��������߁C����������ʌ`���Ƃ͂Ȃ��Ƃ��܂����D�䓡�ق��i2020�j�͓������C���삩��h�E�N�c�~�~�Y�n�[�ƃ��E�X�C�~�~�Y�n�[�ɗގ�����W�{���̏W���C���茧�C�É����C����ыI�ɔ����Y�W�{�������Č����������ʁC�a��ق��i2019�j�Ɠ��l�̗��R���瓯��Ɏ���Ȃ������Ƃ��C�������Y��n�������~�~�Y�n�[���Ə̂���ɂƂǂ߂Ă��܂��D�������C���茧�Y�Ɠ������Y�̕W�{�́C�É����Y�ƋI�ɔ����Y�̕W�{�Ɣ�r���āC��{���̑̒��ɑ��銄�����킸���ɑ傫���X��������C���ꂪ�h�E�N�c�~�~�Y�n�[�ƃ��E�X�C�~�~�Y�n�[�̗L���Ȏ��ʌ`���ƂȂ�\��������Ƃ���܂����i�䓡�ق��C2020�j�D���̂ق��C�؍��ϏB���̗��A�ł́C�`�Ԃ���ꏊ���������̓��A����ɗގ������~�~�Y�n�[�����ނ���������C�������̂悤�ł��i�ϏB��������C2014�j�D
�@�b�����m���ɖ߂��ƁCਉƁE�����i1976�j���h�E�N�c�~�~�Y�n�[�Ƃ������m���Y�W�{�͒��炭�s���s���ɂȂ��Ă��܂����D���̂��߁C�����搶���֗^���ꂽ���{�Y���ޑ�}�Ӂi���m�e���C1984�j�ł̓h�E�N�c�~�~�Y�n�[�̕��z�͈͂ɍ��m�����܂܂�Ă��܂����C�R�k�J���[���ӓ��{�̒W�����i�g�c�E���ÁC1989�j�Ȃ�тɋ������ł���ё��Łi���m�ق��C1993�C2000�j�ł͌듯��ƌ��ꂽ�̂��܂܂�Ă��܂���D�Ƃ��낪�C2001�N�ɍ��m�����b�h�f�|�^�u�b�N�i�����ҁj����̂��߂̕W�{�����̍ہC��������OB�̍����O�����ɂ���Ă���Ǝv�����W�{�������������甭������CBSKU 56156�Ƃ��ēo�^����܂����D���̂Ƃ��C1976�N�ɓ�������OB�Ōܓ����Z�̋��@�߂��Ă����g�x�ˎO�����̏W�����ƍl�����钷�茧�Y�h�E�N�c�~�~�Y�n�[��2�W�{��������C���킹��BSKU 56157�ƂȂ��Ă��܂��D�����āCBSKU 56156�͂Ƃ��鏊�ɑ݂��o����ăh�E�N�c�~�~�Y�n�[�Ɗm�F���ꂽ���߁C������O�Łi���m�ق��C2013�j�ł͖{�������z�͈͂Ɋ܂܂ꂽ�悤�ł��D�������Ȃ���C���m���Y�͔����I�O�ɓ���ꂽ1�W�{�݂̂ł���C���̐����̔c����`�ԓI�����̐������ۑ�Ƃ���Ă��܂����i����C2015�G�����C2018�G�a��ق��C2019�j�D
�@�����ŁC���������ɂ����Ėԗ��I���O��I�Ȓ������s�����Ƃ���C�h�E�N�c�~�~�Y�n�[�Ȃ������E�X�C�~�~�Y�n�[�ɓ��肳���W�{�����m������9���n����V���Ɋm�F����C���ʎ�ƌ���قǑ��Y����͐�����邱�Ƃ����炩�ɂȂ�܂����i�R��ق��C2023�G�����ق��C2024a, b�C2026�j�D�܂��C������BSKU 56156��BSKU 56157�̕W�{�ɉ����ĐV���ɓ���ꂽ�W�{�Q���������ʁC�a��ق��i2019�j���q�ׂ��悤�Ɋ��������̎��ʌ`���͗L���Ƃ͌������C�䓡�ق��i2020�j����������{���ɂ����鐔�l����̓h�E�N�c�~�~�Y�n�[�����E�X�C�~�~�Y�n�[�̗��킪�܂܂��ƂȂ�܂����͍��ł����i�����ق��C2024a, b�j�D�����āCmtDNA�̕ψقł͍��m���Y�W�{�ɕ����킪�܂܂��\���͎x������܂���ł����i�����ق��C2026�j�D�J�ł͓��A�ɂ���h�E�N�c�~�~�Y�n�[�C�N���ɂ�����E�X�C�~�~�Y�n�[�Ƃ����F��������悤�ł����i�Ⴆ�C�����C�m��w���A��������C2025�j�C���m���Y�W�{�ɂ��Ă�BSKU 56156���h�E�N�c�~�~�Y�n�[�Ƃ��ꂽ���j�I�o�܂����܂��C�͐�Y�ł����Ă��h�E�N�c�~�~�Y�n�[�Ȃ���������܂ޕ\�L��p���Ă��܂��i�����C2018�G�R��ق��C2023�G�����ق��C2024a, b�C2026�j�D
�@���ł���ɂ���C�͐�ɂ�����o���ɊԌ��̑������I���╚�����̉e�����K�v�Ƃ����h�E�N�c�E���E�X�C�~�~�Y�n�[�iLuciogobius cf. albus)�́C�͐�����ǍD�ȏ؋��Ƃ��ďd�v�ȑ��݂ł��iKanagawa et al., 2011�G�a��ق��C2019�G�䓡�ق��C2020�G�����C2022�G�����ق��C2026�G�r���E�����C2026�j�D�����āC���m���ɂ͖{��̌����������n��10������i�����ق��C2026�j�C�S���̉ߔ����߂邱�Ƃ͓��M�����ׂ��ł��傤�D�{�킪���Y����m����́u�������{��v��u�m���u���[�v�Ŗ���y���Ă���C�Ԗ���썪��͖����̏��K�͐��n�Ȃ���C�{�����w�̐����ʼnV�∼�̎p���Z���悤�Ɋ����܂��D�܂��C�{�킪��������͐�̏����Ƃ��ẮC�����̉������ɂ��Ԍ��̖ڋl�܂肪�Ȃ��ȑO�ɁC����̒n�����獻�D�����Ȃ����Ƃ��W���Ă���ƍl�����܂��i�����C�����\�j�D�l���̒n���͖k����̉ƑсE�O�g��сE�����сE�l���\�тɋ敪����܂����C�����тƎl���\�т��\������`���[�g�E����E�D�ⓙ�ɂ́C�̉ƑтƎO�g��т��\�����錋���Њ�E�ԛ��ⓙ�Ɣ�r���čӂ��ɂ�������������܂��D�䂦�ɁC�O�҂𗬂��͐�ɂ͖{�킪�D�ފԌ��ɕx���I���������Ȃ�܂��D�����āC�{�킪���z���鍂�m���Ɠ������̑����m���͑S�Ē����сE�l���\�тɑ����Ă���C�{�B�̊��m���z�͐�̈ʒu�������сE�l���\�т̕��z�ƈ�v���Ă��܂��D ���āC�{��͏t�ɎY������Ƃ̕��e�n���炠��i�����ق��C2019�G�䓡�ق��C2020�G����ق��C2020�j�C�Y���̂��߂ɐꏊ�Ɍ���C���̌�͝ˎ����邱�Ƃ���������Ă��܂��i����ق��C2020�j�D���m���ɂ����Ă��C���̉Ԃ��U��G�߂ɂ͍��F�̃n�[�������Y���ɏW���l�q���ώ@�ł��܂��i�����ق��C2024b�j�D���̉Ԃ͓��{�̐��_�̏ے��Ƃ����Ă��܂����C�������͋��y�̐����̏ے��Ƃ��āC�U���Ă͍炭�Ƃ������̉c�݂����`���Ă����������̂ł��D
�@�y����� ���݂��� �l��͂� ���ʁi�݂Ȃ��j�ɓ��� ��i����j����
���p����
���V���G�D2001�D���A�ɐ��ދ��E�h�E�N�c�~�~�Y�n�[�Cpp. 18-20�D�R�����G�E�R���v�E���X�؋��i�ҁj�����Ύ��R�ّ�1����ʓW�u�n�[�̂Ȃ��܂����v�W������D�������������Ύ��R�فC�i���j�z�V�U�L�O���[�����c�C���c�D
�r������E���� ��D2026�D���E�X�C�~�~�Y�n�[�Cp. 100�D�O�d���_�ѐ��Y���݂ǂ苤�����i�ہi�ҁj�O�d�����b�h�f�[�^�u�b�N2025�|�O�d���̐�ł̂�����̂���쐶�����|6�D���E�W�����ށD�O�d���_�ѐ��Y���݂ǂ苤�����i�ہC�ÁD
���m�E��c���v�E��{����E�r�c�S��D1993�D�n�[���ځCpp. 997-1116�D���V�O���i�ҁj���{�Y���ތ��� �S��̓���D��1�ŁD���C��w�o�ʼn�C�����D
���m�E��{����E�r�c�S��E��c���v�D2000�D�n�[���ځCpp. 1139-1310�D���V�O���i�ҁj���{�Y���ތ��� �S��̓���D��2�ŁD���C��w�o�ʼn�C�����D
���m�E��{����E�r�c�S��E���V���G�D2013�D�n�[���ځCpp. 1347-1608, 2109-2211�D���V�O���i�ҁj���{�Y���ތ��� �S��̓���D��3�ŁD���C��w�o�ʼn�C�`��D
�ϏB��������D2024�D�ϏB���A�ΐ��ɖ��L�^�틛�ސ����D(In Korean) LINK
https://www.jejudomin.co.kr/news/articleView.html?idxno=48520#:~:text=%E2%96%B2%20%EC%9A%A9%EC%B2%9C%EB%8F%99%EA%B5%B4%20%EB%82%B4%EB%B6%80%20%ED%9D%AC%EA%B7%80,%ED%98%95%EC%83%81%20%ED%81%B0%20%EC%B0%A8%EC%9D%B4%EA%B0%80%20%EC%9E%88%EB%8B%A4�i30 Apr. 2026�j�D
���Ê�q�D1963�D�h�E�N�c�~�~�Y�n�[�ɂ��āD�����w�G���C72: 1-5.
���Ê�q�D1979�D�~�~�Y�n�[�ނ̐����j�����o���D�W�����C5: 80-84.
���������Y�D2022�D���E�X�C�~�~�Y�n�[�Cp. 149�D�a�̎R����������������� �����������ێ��R�����i�ҁj�C�ۑS��d�v�Ȃ킩��܂̎��R�|�a�̎R�����b�h�f�[�^�u�b�N�|�m2022 �N�����Łn�D�a�̎R����������������� �����������ێ��R�����C�a�̎R�D
���������Y�E�x�� ���E������u�E���R��イ�D2018�D���q��̕���������̏W���ꂽ�I�ɔ������L�^�̃��E�X�C�~�~�Y�n�[�i�ҍ�������E�n�[�ȁj�D��I�����C60: 178-181.
���������Y�E�x�� ���E������u�E���R��イ�E�c�� ���E��{�N�i�E����Εx�D2019�D�Í��삩��̏W���ꂽ���E�X�C�~�~�Y�n�[�i�ҍ�������F�n�[�ȁj�̋L�^�D��I�����C61: 136-138.
�䓡����E�� ����E������P�D2020�D�������C���삩�瓾��ꂽ�n�������~�~�Y�n�[���iPerciformes: Gobiidae�j�̌`�ԂƐ������D���{�����n���w����C75: 18-24.
��c���r�D1934�D�卪���n��詓����̓������D���{�w�p����C9: 301-304.
Kanagawa, N., T. Itai and H. Senou. 2011. Two new species of freshwater gobies of the genus Luciogobius (Perciformes: Gobiidae) from Japan. Bulletin of the Kanagawa Prefectural Museum, Natural Science, 40: 67-74.
���쒼�K�D2015�D�h�E�N�c�~�~�Y�n�[�Cpp. 116-117�D���Ȏ��R���ǖ쐶�����ۊ�ۑS���i���i�ҁj���b�h�f�[�^�u�b�N2014���{�̐�ł̂�����̂���쐶����4�D���E�W�����ށD���傤�����C�����D
���쒼�K�E�쓈�����E���̍N�O�E�䗲�F�E�a��_��D2020�D���������~�~�Y�n�[�����ރi�K���~�~�Y�n�[�ƃ��E�X�C�~�~�Y�n�[�̐����j�|���牺�ɂ�����e�t���̊O���`�Ԃ̕ω��𒆐S�Ƃ��ā|�D���C���R���C13: 65-78.
������㏼�D1955�D���ނ̌`�Ԃƌ���II�D�菑�X�C�����D1605 pp.
�{�n�B�O�Y�E��ߕ��_�ƁE����M�F�D1976�D���F���{�W�����ސ}�ӁD�S�����V�ŁD�ۈ�ЁC���D462 pp.
����Í��D1972�D�ܓ��x�]���̗n��g���l�����ɂ��ރh�E�N�c�~�~�Y�n�[�D���茧�����w��C3: 1-8�D
���茧�D2022�D���茧���b�h���X�g2022. https://www.pref.nagasaki.jp/uploads/2022/04/1648813265.pdf (1 Apr. 2026)
���� ���E�É����d�q�E��쐳�v�E�J�����F�D1984�D���m���̒W�������D���m�������ʋ��Ƌ����g���A����C���m�D156 pp.
���������E�R�㗳���E�����O���E�����L���D2024a�D���m�����甼���I�Ԃ�ɓ���ꂽ�h�E�N�c�~�~�Y�n�[�̐����Ƃ��̌`�ԓI�����D���m��w���H�w���I�v��7���ʍ���1�� ��115��y�������w����u���v�|�W�D
���������E�R�㗳���E�����O���E�b��ÍW�E�����L���D2024b�D���m���ɂ�����C�h�~�~�Y�n�[��Q�̕��z�E��������ь`�ԓI�E��`�I�����DIchthy, Natural History of Fishes of Japan, 43: 20-37. LINK
���������E�R�㗳���E���i ���E�k �F��E���c �āE�����O���E�b��ÍW�E�����L���D2026�D���m���ɂ�����C�h�~�~�Y�n�[�ƃh�E�N�c�~�~�Y�n�[�ߎ���̒lj��L�^�DAquatic Animals�C2026: AA2026-17. LINK
Regan, C. T. 1940. The fishes of the gobiid genus Luciogobius Gill. Annals and Magazine of Natural History (Series 11), 5: 462-465.
�a��_��E���V���G�E��؎��V�E���쒼�K�E�������l�D2019�D�É����Y�~�~�Y�n�[�����ނ̕��ފw�I�����i�\��j�D���C���R���C12: 29-96.
�����O���D2018�D�h�E�N�c�~�~�Y�n�[�Cp. 110�D���m�����b�h�f�[�^�u�b�N�i�����ҁj�������� �����ψ���i�ҁj���m�����b�h�f�[�^�u�b�N2018 �����ҁD���m���ыƐU���E�����������ہC���m�D
�Ɛߖ�E���� ���D1976�D�C�h�~�~�Y�n�[ Luciogobius pallidus �̐��Ԃɂ��āD���a51�N�x���{���ފw�w��N��u���v�|�W�D���{���ފw��C�����D28 pp.
�����C�m��w���A��������D2025�D�}�^ �~�~�Y�n�[���Q�D�����C�m��w���A��������C�����D
Tomiyama, I. 1936. Gobiidae of Japan. Japanese Journal of Zoology, 7: 37-112.
�R�㗳���E���������E�����O���E�����L���D2023�D���m���ɂ�����n�����~�~�Y�n�[�����ނ̕��z�Ɛ����D���m��w���H�w���I�v��6���ʍ���1���C��114��y�������w����u���v�|�W�D
�R�����G�D2014�D�h�E�N�c�~�~�Y�n�[�Cp. 82�D�������܂˃��b�h�f�[�^�u�b�N�i2014�����ҁj�D�������������������R���ہC���]�D
�g�c���j�E���Ê�q�D1989�D�C�h�~�~�Y�n�[�Cpp. 628-629�D��ߕ��_�ƁE����M�F�i�ҁj�R�k�J���[���� ���{�̒W�����D�R�ƌk�J�ЁC�����D
�ʐ^�W�{�FBSKU 160909�C27.9 mm SL�C���m�����|�S���m�� �썪��C2026�N4��29���C�̏W�E�B�e�F���c �āE���������D
�i���������j
2026�N5���̋�
![]()

�����n�^Aulacocephalus temmincki Bleeker, 1854�i�X�Y�L�ڃn�^�ȁj
�@���������C���������������̋��ȂǂƂ����C��ʓI�ɂ͂܂����邱�Ƃ̂Ȃ��悤�ȃT�C�g�����ɗ���l�ԂƂ����̂́C�����Ă��d�x�̋����^�N������ɏ�������̂ł���Ƒ��ꂪ���܂��Ă���D�L���������Ă���{�l�܂߁C���D���Ƃ����l��͂��̐��ɂ��܂�Ƃ��āC�����������͂��ꂪSNS�Ȃǂɂ������������̂�����Ȃ��̂��ƃ^�`�������D�悭������g����������Ă���l�Ԃ́C���Ɍ��I�ł���h�Ƃ����ӌ��̍������E���W�߂Ă�����X�ł���Ə�X�����Ă���D
�@���āC����ȋ�����D���ȉ�X�͒��x�̍��͂���C�F��������D���ȋ��Ƃ����̂����݂���D�D���ȗ��R�Ƃ����̂͐獷���ʂ����i�����Ƃ����������Ƃ��C����Ă��Ă����������Ƃ��C�Q�[���t�B�b�V���Ƃ��ėD�G�Ƃ��c�j�C�l�I�ɂ͐F�����d�v�ȃt�@�N�^�[�ł���ƋC�Â����̂ł���D���Ȃ킿�C�������D���Ȃ̂ł���D
�@�������Ƃ��C���ɂƂ��Ă̐F�Ƃ����͎̂��͒��ׂĂ݂�ƂȂ��Ȃ������[���D�Ƃ����̂����ݒm���Ă���F�̋��̂قƂ�ǂ́C�����Ɍ����g��������h���Ȃ̂ł���D���w�����ȓǎ҂̊F����Ȃ�s���Ƃ����Ǝv���邪�C����͂�����\���F�Ƃ������̂ł���D�\���F�Ƃ́C�����̂̐������قȂ���̔g���ɋN�����āC���̂ӂ�܂����ω����邱�ƂɗR���������F���ۂ̑��̂ł���i�؉��ق��C2002�j�D���ۂ͂���ȐF�͂��Ă��Ȃ����C���˂��������\�ʂ̍\���ɂ���Ĕ��ˁE���E��܂��ĔF�������F�ʂ̂��Ƃł���ƁC���͂����F�����Ă���D���ȊO���ƃ����t�H�`���E���̒��Ԃ��L���ł���D���̐����Y��ō������Ȓ����C���̐l�͂Ȃ�ƂȂ��C���[�W�ł���̂ł͂Ȃ����낤���D�����C�������g�{���ɐ��h���Ƃ����̂��m���Ă���C�j�V�L�e�O���̒��ԁi������}���_�����t�B�b�V���̂��Ɓj������ɊY������i�哇�C2016�j�D���̃O���[�v�̂݁C�F�̐F�f��L���邱�Ƃ��m���Ă���D��������킩��̂́C�ǂ���玩�R�E�ōs���Ă���i���̃��[�X�ɂ����āC�{���̐F�i�F�F�f�j��L����̂͂ǂ������ɏd���R�X�g��������C�Ƃ������Ƃł���D���̐��ɂ͐F�́i������X���F������j��������Ȃ�ɑ��݂���Ƃ������Ƃ́C�����炭�F�͉�������ɑ��ēK���I�ł���C���̂��߂Ɋl������o�H�Ƃ��Ă͍\���F�̂ق�����R�X�g�ł������Ƃ������Ƃ��낤�D���ꂩ�C������������قƂ�ǂ̋��ނŐF�̐F�f����ɓ���邱�Ƃ�����Ȃ�悤�ȃ{�g���l�b�N�����݂��Ă���̂�������Ȃ��c�ƁC�����܂ł��Ǝv�������Ƃ𗅗Ă݂����C�悭�悭�l����ƁC�͂����č\���F�Ƃ����łȂ��F��{���ɒP���ɓ����F�Ƃ��ĂƂ炦�Ă����̂��H�Ƃ����C�ɂ��Ȃ��Ă����D���ɓI�ɂ����C�F�Ƃ����̂́C�܂����������Đ�ΓI�Ȃ��̂ł͂Ȃ��C�Ђǂ���ϓI�Ȃ��̂ł���D�Ⴆ�Ύ��ɂƂ��Ă̐ԐF�����Ȃ��ɂƂ��Ă̐ԐF�Ɗ����ɍ��v���Ă��邱�Ƃ͌����ł��Ȃ��킯�ŁC���g�ɂƂ��Ă̐F�ɑ���F���Ƃ����̂͌X�Ɉς˂��Ă���D�܂�C��X�̓}���_�����t�B�b�V�������̋��������F�̋����Ǝv���Ă��邪�C���ɂƂ��ė��҂̐F�͑S���Ⴄ���̂Ƃ��ĔF�������̂ł͂Ȃ����낤���H���̊�̍\���Ȃǎ�ɂ���đ傫���قȂ�ł��낤���Ƃ͑z���ɓ�Ȃ��D���Ȃ��Ƃ���X�l�ԂƂ͈قȂ邾�낤�D��������ƁC����ʓI�ɍL���K���I�Ȃ͍̂\���F�̐F�Ƃ������ƂɂȂ��Ă���D���ɑ���F�̋c�_�ɂ́C���̂悤�Ȗ�肪��ɕt���܂Ƃ��D�N����l�C�w�W�ɂȂ蓾���ΓI�Ȑ_�l��������Е����������̂����C���ǂ̂Ƃ��닛�ɂƂ��Ă̐F�ʂƂ����͉̂�X�ɂ͗����̋y�ʔ��e�Ȃ̂�������Ȃ��D
�@�����Ԙb������Ă��܂������C���炽�߂Ď��̍D�݂̘b������ƁC����܂łɂ��������������F�ɏo����Ƃ��ł����D�Ăɋ����Ă���ɂ���Ă���C�J��ނ��Ă����Ƃ��ɖԂŋd�����o�V���E�J�W�L�̎ዛ��C��u�Ԃɓ������g�r�E�I�̗C�ނ肽�ẴE���C����Ί_���̃e�[�u���T���S�̉��Ō����i�C�g�J���[�̃c�`�z�[���Ȃǂ��v���o����邪�C�Ȃɂ��C��Ԋ��������̂�����̖{��ɂ����郋���n�^Aulacocephalus temmincki Bleeker, 1854�ł���D�����n�^�C�����ŏ����Ɨڗ��H���ł���D�Ȃ�Ƃ܂��C�������O�ł��낤���D���ۂɂ݂���ڗ��F�Ȃ̂�����C���҂𗠐�Ȃ����ł���D�ǂ����Ă��C���O�̂��ɑS�����Ȃ��A�I�n�^�Ƃ̍���Ɋ����Ă��܂��D���̋��������n�^�ƌĂюn�߂��l���Ɋ��ӂ̋C������`�������Ƃ���ł���D�ʐ^�̌́iBSKU 135048�j�͎��˖����̐��[ 100 -150 m�ŕߊl���ꂽ���̂ŁC�������ƈꏏ�ɒނ蒲���ɎQ�����Ă��������ق̃X�^�b�t�̈�l����ՓI�ɒނ�グ����C�ł���D������������̊����́C����͂������������̂ŁC�����n�^���̂͂����܂łƂĂ��������킯�ł͂Ȃ����̂́C���߂Č������̃����n�^���������߁C���̎��̎��͕������������̂��̂悤�ɂ͂��Ⴂ�ł����̂ł���D�K���s�K�������̕ω��ɂ��_���[�W�őh��������Ȃ��������߁C�W�{�Ƃ��Č������ɏ����Ă����������D���肪�����b�ł���D
�@�����n�^�̓����I�Ȑ��Ԃ̈�ɁC�h����^����Ɣ畆����O���~�X�`���ƌĂ��S�t�ł���o����Ƃ������̂��m���Ă���i�����C1987�j�D���̔S�t�͍����S���������C���͂̐����A���悤�ɂ݂��邽�߉p�ꌗ�ł�Soap fish�i�Ό����j�ƌĂ�Ă���D���͂̋��͎���ł��܂��炵���D�Ȃ��C���l�̓����������́C���{�����ɂ����Ă̓����n�^�ȊO�ɂ̓k�m�T���V�C�L�n�b�\�N�C�A�S�n�^�C���~�X�Y�L���m���Ă��邪�C��������ʑ��ł���C���̓��������L�h���`���Ȃ̂��C����Ƃ����n���I�Ɋl�����ꂽ�̂��͋c�_�̗]�n������D�悭����b�Ƃ��āC�������Ƀk�m�T���V�i��L��Soap fish�̒��ł͈�Ԗڂɕt���@������C���炵�₷���j�����Ă�����C�ł��o���Ă��܂��ăk�m�T���V����Ƃ��������̋����S�ł��Ă��܂����Ƃ����̂�����D�����̐��ʂ�������ǂ��ɂ��Ȃ�悤�����C���̎��v���e�C���X�L�}�[�i�C��������ɗp������C�A�̗͂ŗL�@��������t�B���^�[�̈��j����͂������ʂ̖A���o�Ă���悤�ł���D�������C����Soap fish�Ƃ����ď̂Ƃ����̂͂悭�悭�l�������a���̂��閼�O�ł���D�A������Ό��Ƃ����̂́C����͂������낤�Ǝv��Ȃ��ł��Ȃ����C�A���Ƃ����͔̂S�x�̍����t�̂���C���������ޏ�Ԃ̂��Ƃł���D�ł���C�{���ނ炪�������鐅���ɂ͂��̂悤�ȋC�̂͑��݂��Ȃ��킯�ŁC�O���h���ɑ��鉞����[�I�ɕ\�������̂Ƃ�����肩�́C�l�Ԃɂ��h���i�܂�C��C���Ɉ�������o�����Ƃ������Ɓj�ɑ��Ă݂̂̉�����\�������̂Ȃ̂ł͂Ȃ����낤���D���ɂ���CSoap fish�͊O�I�h���ɑ��鉞���Ƃ��ĔS�t�ł���o����Ƃ������Ƃ͊m���炵���D�������Ȃ���C���̃g���K�[���\�c�ꗍ���ɊO�I�h���Ƃ��Ă܂Ƃ߂Ă��܂��̂́C��≡�\�̂悤�ȋC�����Ă��܂��̂ł���D
�@���āC�D�̏�ŏ��߂ă����n�^����ɓ���Ă����x�̓����̎��́C�ӂƂ��邱�ƂɋC�������D����́C�̕\���為��S�t���o�Ă��Ȃ��̂ł���D���n�]�ʂ�ɂ����C�O�I�h���i�ނ�グ��ȂNJO�I�h���ȊO�̉����ł��Ȃ����낤�j�ɂ��S�t�ł���o����̂ł͂Ȃ������̂��H�Ȃ����҂͂���ł���D���͕W�{�ɂ���Ƃ��C������x�S�t������Ă��珈�����s���̂����C���̎��������܂ŔS�t�����ʑ����킯�ł͂Ȃ������D��������n�^���S�t���o���^�C�~���O�͂����ƌ���I�Ȃ̂��낤���D�����v���C�����n�^�����x���ނ������Ƃ̂���F�l�i�����ɂ���ƈӖ����킩��Ȃ����C���ۂɑ��݂���j�ɕ����Ă݂�ƁC������ł͒ނ����Ƃ��ɔS�t���o���Ă����炵���D���悢��킩��Ȃ��Ȃ��Ă����D����̌̂����܂��܂������̂�������Ȃ����C�����n�^���Ƃ�Soap fish�̓ł̕��o�ɂ͂�茵���ȏ���������̂�������Ȃ��D
�@�����܂Ń����n�^�Ɋւ��Ă��Əq�ׂĂ������C��͂肱��قǂ������������C���ފw�҂ł���O�Ɏ��牮�ł���Ƃ���̎��Ƃ��ẮC�ǂ����Ă�����~���h������Ă��܂��D���Ј�ĂĂ݂������̂ł���D�����n�^���̂͊Ϗ܋��Ƃ��Ă͂���Ȃ�ɗ��ʂ��Ă��邽�߁C��낤�Ƃ���w�����邱�Ƃ��\�ł���c���C���͂��̒l�i�ł���D����������ƁC�S��7cm�̌̂�298000�~�Ŕ̔�����Ă����i�I�j�D�����āC�撣��Β��Â̌y�����Ԃ�������悤�Ȓl�i�Ȃ̂ł���D�ǂ��l���Ă��n�R��w���������ɂ̓n�[�h�����������鉿�i�ݒ�ł���D�܂��C�[��ɐ������Ă��āC�_���ĕ߂邱�Ƃ��ł��Ȃ��āC���Ϗ܋������̉f����F�ʂł��邩�獂���͎̂d���Ȃ��̂�������Ȃ��D���ł�������Ȃ����낤���D
�@���݁C���̎���ɂ͋��60cm�I�[�o�[�t���[�������ݒu����Ă���D�A�N�A���X�g���邠��̈�ŁC��̐����𗧂��グ�ĉĂ���Ƃ�����Ԋy�����Ƃ����̂�����D����́C�����炭���̐����łǂ�ȋ������������Ƃ����z�������Ă���̂��������y�����Ƃ������Ƃ��Ɖ��߂��Ă���D�܂�Ƃ���C���͖�Ȗ�ȋ���ۂ̐������Ȃ��߂ă����n�^�����炷��z�����Ƃ��ϑz�����Ă���킯�ł���D�w������̂͌����I�ł͂Ȃ��C�����Ȃ�Ύ����Œނ��ĕߊl���邵���Ȃ��D�������ނ邾���ł͂Ȃ��C�������Ԃ悭�h��������K�v������D���Ƃǂ������\�������̂��H�Ǝv��Ȃ��ł��Ȃ����C����ł��ǂ��ɂ����Ă�낤�Ƃ����Ǝv�������Ă���D�������Ɏ���܂ł����������Ƃ��āC���������̒��œł��o���Ď��ł��Ă�����H���̎��́C�܂��ނ��Ă��邵���Ȃ��̂��낤�D���ꂷ����ϑz�����C���Ȃ��Ƃ������n�^���炵������f�ɂ܂�Ȃ��̂͊ԈႢ�Ȃ����낤�D
�@����͍D���ȋ����o���_�Ƃ��ĕ��͂����������C�D���ȋ��Ƃ����̂͂����܂ł��l�Ԃ̈���I�Ȋ���ł����Ȃ��C���ɂƂ��Ă͂ނ���^�t�̊���قƂ�ǂł��낤���Ƃ͑z���ɓ�Ȃ��D�������Ȃ���C��X�l�Ԃ̃G�S�����܂������̂ŁC���͂��̕Ўv��������������Ƒ����Ă��������ł���D
���p����
�����@���D1987�D���F���ޑ嚤�ӁD�k���فC�����D1029 pp�D
Bleeker, P. 1855. Nieuwe nalezingen op de ichthyologie van Japan. Verhandelingen van het Bataviaasch Genootschap van Kunsten en Wetenschappen, 26 (art. 4): 1-132, pls. 1-8.
�؉��C��E�g���L��E����N�T�D2002�D���R�E�̍\���F�̎d�g�݁D�F�ދ��, 75 (10): 493-499�D
�哇�͎q�D2016�D���̑̐F�Ƃ��̕ω��F���J�j�Y���ƍs���w�I�Ӌ`�D�F�ދ��, 89 (6): 178-183�D
�ʐ^�W�{�FBSKU 135048�C145.2 mm SL�C���m�����˖����C���[ 100-115 m�C�ނ�C�C���C2024�N1��9���D�̏W�ҁF���c�D��i�����C�m��SATOUMI�j�D
�i�F�،d��j
2026�N4���̋�
![]()

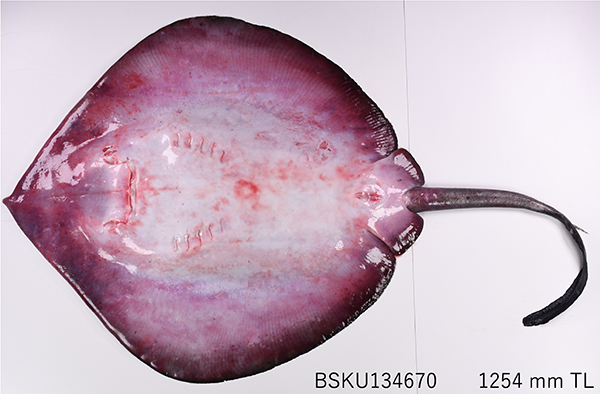
�E�X�G�C Plesiobatis daviesi (Wallace, 1967)�i�G�C�ڃE�X�G�C�ȁj
�@����́C�[�C�ɐ������鋐��ȃG�C�C�E�X�G�CPlesiobatis daviesi �����Љ�܂��D �E�X�G�C�� Plesiobatis Nishida, 1990�́C������̃E�X�G�C1��݂̂��܂ރO���[�v�ł��iFricke et al., 2026�j�D
�@�{��̓C���h�E�����m�̓�A�t���J����n���C�����ɂ����čL�����z���Ă��܂��iNelson et al., 2016�GSeret and Last, 2016�j�D�����ł́C���J�i1982�j�ɂ���B-�p���I�C�䂩�珉�߂ĕ���C���̌�ɂ͈�錧�����t���َR�p�C���͘p�C�O�d���C���m���C���茧�C�����C���ꌧ�C����M��C�~�Ȃǂ���L�^����Ă��܂��i���V�ق��C2013�G��R�ق��C2013�G�����ق��C2015�G�{���C2026�j�D���m���ł́C�y���p���ł̐��Y�����������D�������ۂɂ��g���[�������œ���ꂽ�W�{����ɁC�O�V�E�����i2014�j�ɂ�菉�߂ĕ���܂����D
�@�E�X�G�C�́C�ҍE��5�ŁC������������[�����C�w�h���Ȃ��C������1�C2�{����C���h�������C�@�E��ق����킸���ӂ��Ђ���ɂȂ邱�ƂŃG�C�ڂ̑��ȂƎ��ʂ��\�ł��D�i���V�ق��C2013�G��R�ق��C2013�GSeeret and Last, 2016�j�D�{��̑̐F�͔w�ʂ��D�F�܂��͐Ԋ��F�ŁC�������Ê��F�C�����Ĕ��h�����F�C���ʂ����F�Ŏ��͂����Â�������邱�Ƃ������ł��D �{��͓�A�t���J�œ���ꂽ�W�{�Ɋ�Â��L�ڂ���C�����͂��̒������h�𗝗R�� Urotrygon �ɕ��ނ���Ă��܂����i�]���ق��C2010�G��R�ق��C2013�GFricke at al., 2026�j�D�������CNishida�i1990�j�͌`�Ԍ`���Ɋ�Â��g�r�G�C���ڂ̕���w�I��͂ɂ��C�E�X�G�C���g�r�G�C���ڂ̋��ʑc�悩�瑁���i�K�ŕ����ꂽ�Ǝ��̌n���ł���C�n�����̊�Ɉʒu���鉼���������܂����D�]����Urotrygon�ɃE�X�G�C���܂߂邱�Ƃ́C���ފw��́u�P�n�����v�Ȃ����߁C�Ɨ������E�X�G�C�ȁiPlesiobatididae�j����уE�X�G�C���iPlesiobatis�j��V�݂��Ă��܂��iNishida, 1990�j�D
�@������ Plesiobatis �́C���̌��n�I�Ȍn���I�ʒu�f���C�M���V����Łu���n�I�ȁv���Ӗ����� plesio �Ɓu�G�C�v���Ӗ����� batis ��g�ݍ��킹�����̂ł��iNishida, 1990�G�O�V�C2024�j�D�Ȃ��C�포���� daviesi �́C��A�t���J�̊C�m�������iOceanographic Research Institute�j�����ł������G�C�ނ̌����� David H. Davies ���ւ̌����ł��i���V�E�����C2015�j�D�E�X�G�C�͍ő�S�� 270 cm�C�̏d 42 kg �ɒB�����^��ŁC�I�X�ł͑S�� 130-170 cm�C���X�ł�189-200 cm�Ő��n���܂��D��̂̎q�{���番�傳���^���p�N���Ǝ��b��L�x�Ɋ܂ގq�{���t����h�{��⋋����g�D�h�{�^�i�q�{�~���N�^�C�v�j�ِ̑��ŁC�o�����̑S���͖� 50 cm �ɋy�т܂��iSeret and Last, 2016�j�D �E�X�G�C�͉p���� �gDeepwater stingray�h �ƌĂ��ʂ�C�����ɐ��[ 275-680 m�̑嗤�I��嗤�Ζʂɐ������C���^���ނ�b�k�ށC�����ނ�ߐH���Ă��܂��iEbert et al., 2002�GSeret and Last, 2016�j�D�[�C���̃C���[�W�������ł����C��C��ł̋L�^������C���U���r�[�N�ł͐��[ 44 m����C���茧�ł͐��[�� 25 m�̒�u�Ԃŋ��l����܂����iWhite et al., 2006�G�����ق��C2015�j�D���̐��Ԃɂ́C���܂������̓䂪�c����Ă��܂��D
�@����̎ʐ^�́C���m�s��������`�̑��J��ԋ��Ŏi�ۂɂ�葫�����������̐��[ 340 m�ō��l���ꂽ�̂ł��D�E�X�G�C�͐�[���s��2�{�̔����ɓł������Ƃ���C���l�����ƑD��ł����ɔ��h���Ɛ�̂Ă��Ă��܂��܂��D�������C�ʐ^�̂͂��̓��Ɏi�ۂ֏�D�����{�������̌F�،d�펁�i���� M2�j�ɂ��C�����Ɣ��h�����銮�S�ȏ�Ԃō̏W����܂����D�����C�܂��������z���O���������ɂ͒m��R������܂��C��y���킭�u���傷���ĕW�{�쐬�ɂ͎��ʂقNj�J�����v�Ƃ̂��ƁD�[�C�̈Èł��猻�ꂽ���̋���Ȏ��O�ɁC�i�������y�����̎p���ڂɕ����Ԃ悤�ł��D
���p����
���V���G�E�R���֎q�E�������ȁD2013�D�E�X�G�C�ȁD���V�O���i�ҁj�Cpp. 217, 1773. ���{�Y���ތ��� �S��̓���C��O�ŁD���C��w�o�ʼn�C�`��D
�����b���q�E�o���^�R���E��v�ۏ����D2015�D���茧�ɂ�����E�X�G�C�iPlesiobatis daviesi�j�̏��L�^�D���茧�����w��C(77): 10-12.
Ebert, D. A., P. D. Coeley and L. J. V. Comoagno. 2002. First records of the longnose spiny dogfish Squalus blainvillei (Squalidae) and the deep-water stingray Plesiobatis daviesi (Urolophidae) from South African waters. African Journal of Marine Science, 24: 355-357.
Fricke, R., W. N. Eschmeyer and R. Van der Laan. 2026. Catalog of fishes: genera, species, reference. (27 April 2026)
�ؑ����u�D2025�D�E�X�G�C�D�ؑ����u�E���ؑ�n�i�ҁj�Cp. 38. �O�d���̋��ސ}�ӁC�ؑ����u�C�ɐ��D
�O�V �ɁD2024�D�E�X�G�C�ȁD���{ ���E���V�O���i�ҁj�Cp. 26. ���{�̐[�C���}�ӁD�R�ƌk�J�ЁC�����D
�O�V �ɁE�����L���D2014�D�W�{�Ɋ�Â������m���Y�G�C�ނ̃`�F�b�N���X�g�D���җތ������C(50): 1-16.
�]�����F�E�Ό� ���D2010�D��錧�������ō̏W���ꂽ�k���̃E�X�G�C�D���җތ������C(46): 38-40.
�{���_�V�D2026�D���{�Y���ޑS��ژ^�D����܂łɋL�^���ꂽ���{�Y���ޑS��̌��݂̕W���a���Ɗw���DOnline ver. 38. (27 April 2026)
���V�O���E�����`�G�D2015�D���{�Y���ޑS��̊w�� �ꌹ�Ɖ���D���C��w�o�ŕ��C�`��Dxvi + 372 pp.
���J��G�D1982�D�E�X�G�C�i�V�́j�D���� ���E�M�v�E�O�J���v�i�ҁj�Cpp. 56-57. ��B�|�p���I�C��Ȃ�тɓy���p�̋��ށD���{���Y�����ی싦��C�����D
Nelson, J. S., T. C. Grande and M. V. H. Wilson. 2016. Fishes of the world. 5th ed. John Wiley and Sons, Hoboken. xli + 707 pp.
Nishida, K. 1990. Phylogeny of the suborder Myliobatioidei. Memoirs of the Faculty of Fisheries, Hokkaido University, 37: 1-108.
��R���v�E���\ �G�E���{ �t�D2013�D���͘p�ɂ�����E�X�G�C�i�G�C�ڃE�X�G�C�ȁj�̋L�^�ɂ��āD�_�ސ쎩�R�������C(34): 77-82.
Seret, B. and P. R. Last. 2016. Giant Stinggarees Family Plesiobatidae, Pages 674-675 in P. R. Last, W. T. White, M. R. de Carvalho, B. Seret, M. F. W. Stehmann and G. J. P. Naylor, eds. Ray of the world. CSIRO Publishing, Clayton.
White, W. T., P. M. Kyne and H. Holtzhausen. 2006. Plesiobatis daviesi. The IUCN Red List of threatened species, 2006: e.T60111A12303835. http://dx.doi.org/10.2305/IUCN.UK.2006.RLTS.T60111A12303835.en (27 April 2026)
�ʐ^�W�{�FBSKU 134670, �S��1254 mm�C�y���p�C�������������C���[ 340 m�C���m�s��������`�C�i�ہC���J��ԁC�̏W�� �F�،d��C2023�N10��23���D
�i�D�R���m�j
2026�N3���̋�

�C�h�~�~�Y�n�[ Luciogobius pallidus Regan, 1940 �i�n�[�ڃn�[�ȁj
�@�n�[�ȃ~�~�Y�n�[�� Luciogobius Gill, 1859�̃C�h�~�~�Y�n�[�́C�̂̐F�f�������Ⴊ�މ������n�������̋��ނł���C�w�h���h�𐔂�10-13�C�\�h���h�𐔂�10-13�C���Ғō�����34-37�C���ō�����18-19�Ȃǂ̓_���W���Ƃ���܂��i�a��ق��C2019�j�DL. pallidus ��Tomiyama (1936) ���~�~�Y�n�[ L. guttatus guttatus �̌̕ψقƂ��Ď������C�x����˂��瓾��ꂽ�W�{�i�̂̐F�f�������C�Ⴊ�މ��k�����Ĕ牺�ɖ��v������������j�̑}�}����ɁCRegan (1940) �ɂ���ĐV��L�ڂ���C�����i1955�j�ɂ���ăC�h�~�~�Y�n�[�̘a��������܂����D���������āCTomiyama (1936) ���p�����a�̎R�������S��V���C���Q���F�a���s�C����э��m�������S�O�葺�Y�̌v3�W�{���{��̃V���^�C�v�ƂȂ�܂��i�a��ق��C2019�j�D
�@�����͈ꕔ�n��̈�˂���H�ɓ�����ɂ߂Ē��������Ƃ���Ă��܂������C����������ਉƐߖ펁�Ɖ��� �������ɂ�鍂�m���V����ł̒����ɂ��C�͏����@�邱�Ƃɂ���ėe�Ղɕ����̕W�{��������ꍇ�����邱�Ƃ���������܂����i�ƁE�����C1976�G���ÁC1979�j�D�Ȍ�C�ˑR�H����Ƃ͂���Ȃ�����i���ȁC2020�j�C�{��̕��z��͐V��������ш�錧���玭�������܂ł̊e�n�C����ɂ͊؍��i���ϓ��C�ϏB���j�ɂ܂ōL�����Ă��܂��iKim, 2012�G���m�ق��C2013�G�a��ق��C2019�G���}�ق��C2020�G�R���ق��C2021�GOhmyNews�C2024�G���F�ق��C2025�j�D
�@���m���ł͖{��ɂ��āC�͐���̈����ɋɂ߂Ďキ�C�V���삪�S���I�ɋM�d�ȑ��Y�n�ł���Ƃ��ꂽ���ƂȂǂ���C��Ŋ뜜�T�ނ���э��m���w��쐶���A���Ƃ��Ă��܂��i�����C2002�G�����C2018�j�D�������C���m���ł͐V����ȊO�̐����͂悭�킩���Ă��Ȃ��̂�����ł����i�����C2018�j�D�����ŁC���������ɂ����Ėԗ��I���O��I�Ȓ������s�����Ƃ���C������40���̐��n�ɐ������C�V����Ɠ����ȏ�ɑ��Y����͐���������邱�Ƃ����炩�ɂȂ�܂����i�R��ق��C2023�G�����ق��C2024�C2026�j�D�n���������ނ͖Ԃ�ނ�Ƃ������ʏ�̋��ޑ������ł͔����ł��Ȃ����Ƃ���C�������ߏ��]������Ă����ƍl�����܂��i���E���R�C2014�G�R���ق��C2021�G�����ق��C2024�j�D
�@���˂Ă��C�C�h�~�~�Y�n�[�ɂ͕�����̍��݂���������Ă��܂����i�Ⴆ�C�g�c�ق��C2006�G�a��ق��C2019�G�����ق��C2021�G�����ق��C2024�j�D���ł��C�a��ق��i2019�j�̓V���^�C�v3�W�{����2�^��F�߁i���m���Y�W�{�Ƙa�̎R������ш��Q���Y�W�{�j�C�����ق��i2021�j�͒W����̕W�{�Q�i�W���^�j�ƋD����̕W�{�Q�i�D���^�j�̊ԂŌ`�ԓI���ق�����Ƃ��܂����D�������C�W���^�ƋD���^�́C�����ق��i2021�j���������Ғō����Ȃǂ̌v���v���l�͕W�{���𑝂₷�ƕψٕ����d�����Ď��ʌ`���Ƃ͂Ȃ肦�܂���i�����ق��C2024�C2026�G�����C2024�G���F�ق��C2025�j�D����C���N���ɂ͎��̌`�ԂƐF�ʂ̍��ق��猩�����邱�Ƃ��ł��܂��i�����ق��C2024�C2026�G�����C�m��w���A��������C2025�j�D�W���^�͓����w�ʂ̗֊s���̎������ɂ��ג����p�ێl�p�`�ŁC��Ƌ��h���̒��Ԃ̂�����œ������ő�i�� �D���^�F�������`�ŁC��Ƌ��h���̒��ԕt�߂œ����̍ő�j�C���|���ӂ̊זv���[���C���̌���ł͍��E����◲�N���Ē����͕��R �i�� �זv���C���̌���ł͍��E�����N���Ē����͖c���j�C���|����̍��E�̗��N�ɔ��݂�тт镔�������� �i�� �Ȃ��j�D�����āC�W���^�Ɣ�ׂċD���^�͑̐F���Z���i�����ق��C2024�C2026; Ito et al., 2025�j�C�����i2002�j�̊`�F��Kim (2012)�̎�g�F�Ƃ����\�����悭�����܂��D�������C�{�����ނ̓z���}�����Œ�ɂ���Č`�ԂƐF�ʂ��傫���ω����邱�Ƃ��m���C�{�����O�ł͂���܂���i�a��ق��C2019�G�R���ق��C2021�G�䓡�ق��C2024�G�����ق��C2026�j�D�����Ƃ��C���������̎R�㗳�����̌����ł́C�_�o���W�ɂ��`�ԓI���ق����o���Ă��܂��i�����ق��C2026�j�D
�@�܂��C���҂�mtDNA�̌n����͂ɂ��C��`�I�ɖ��Ăɕ������Ă��邱�Ƃ�������Ă��܂��i�����ق��C2024�C2026�G�䓡�ق��C2024; Ito et al., 2025�j�D����ɁC�C�h�~�~�Y�n�[�͏H����~�ɎY������Ƃ���܂����i�����C2002�G�����C2024�j�C�W���^�̕����̂͏t����Ăɓ����Ă���C���������̑����͋D���^�̒m���ƍl�����邽�߁C���҂͎Y�������قȂ�\��������܂��i�����ق��C 2026�j�D�{�e��BSKU 160132��3���̏W�Ȃ�����ɗ����O�����瓧���Č����邽�߁C�t�Y���𗠕t����W�{�ł��D�����āC�C�h�~�~�Y�n�[�ɂ͓����Ƀz���^�C�v�ȊO�̕W�{�������Ă��Ȃ��l�����~�~�Y�n�[ L. dormitoris Shiogaki and Dotsu, 1976 ������܂��D�g�c�ق��i2006�j��Ito et al.�i2025�j�́C�`�ԓI�������l�����~�~�Y�n�[�ɗގ�����W�{���̏W�������CmtDNA���͂̌��ʂɃC�h�~�~�Y�n�[�Ƃ̍��ق͂Ȃ������Ƃ��܂����D�����ق��i2024�j�͌`�ԂƐF�ʂ̓����Ȃǂ���C�g�c�ق��i2006�j�̕W�{�͋D���^�Ɠ���̉\��������Ƃ��Ă��܂��DIto et al. (2025)�́C�������̕W�{���C�h�~�~�Y�n�[�̒��ł��D���^�̈�`�I�ψقɊ��S�ɓ���ꂽ�Ƃ��Ă��܂��D�܂�C�l�����~�~�Y�n�[�ƃC�h�~�~�Y�n�[�D���^�͎���ψق̉\��������܂��iIto et al., 2025�j�D���������̒؈䏮�����̌����ɂ��C���m�������10�N�ȏ�O�Ƀl�����~�~�Y�n�[�ɗގ�����W�{�����������Ă����悤�ł����C���Y�W�{�Q�͌��ݍs�����m�F�ł��܂���D
�@�Ȃ��C�O�q�̃C�h�~�~�Y�n�[�̕��z�L�^�͂قƂ�ǂ��D���^�ƍl�����C�W���^�͍��m����17���n�ƁC���O�ł͐É����C�O�d���C�a�̎R���C�������C�{�茧�C����ѓ������̊e1-3���n����m����݂̂ł��i�����ق��C2021�G�����ق��C2024�C2026�G�䓡�ق��C2024�G�����C�m��w���A��������C2025�G�����ق��C2026�G�r���E�����C2026�j�D�W���^�͋D���^�ɔ�ׂĒn�����ւ̈ˑ��x���������Ƃ���������邽�߁i�����ق��C2024�C2026�j�C�{���ɒn�������L�x�Ȑ���ȉ͐삪�����؋��Ƃ��ďd�v�ȑ��݂Ƃ�����ł��傤�D �Ō�ɁC�����ق��i2024�C2026�j�̏㈲�Ɏ���܂ŁC�ߍ��Ȓ����ƌ����ɂ����͂��������������̏����C�����͂��������������҂̕��X�C��������ӂ��Ă����������搶�ւ̊��ӂ̈ӂ��L���ĕM�����������Ǝv���܂��D
���p����
���m�E��{����E�r�c�S��E���V���G�D2013�D�n�[���ځCpp. 1347-1608, 2109-2211�D���V�O���i�ҁj���{�Y���ތ��� �S��̓���D��3�ŁD���C��w�o�ʼn�C�`��D
�r������E���� ��D2026�D�C�h�~�~�Y�n�[�Cp. 103�D�O�d���_�ѐ��Y���݂ǂ苤�����i�ہi�ҁj�O�d�����b�h�f�[�^�u�b�N2025�|�O�d���̐�ł̂�����̂���쐶�����|6�D���E�W�����ށD�O�d���_�ѐ��Y���݂ǂ苤�����i�ہC�ÁD
���Ê�q�D1979�D�~�~�Y�n�[�ނ̐����j�����o���D�W����, 5: 80-84.
�����L���D2018�D�C�h�~�~�Y�n�[�Cp. 99�D���m�����b�h�f�[�^�u�b�N�i�����ҁj�������� �����ψ���i�ҁj���m�����b�h�f�[�^�u�b�N2018 �����ҁD���m���ыƐU���E�����������ہC���m�D
���������Y�D2024�D�a�̎R���I�̐�͌���ɂ�����C�h�~�~�Y�n�[�̐���������ѐ��B�B�w���C�̏d�ʎw���C�����Ǔ��e���̋G�ߕω��D��I�����C66: 3-8.
�� ����E���R���F�D2014�D�{�B�E�l���E��B�̉͌������ɐ�������n�[�ށD���ފw�G���C61: 105-109.
�䓡����E�� ����E���䏲�q�D2024�D���������Ð삩��̏W���ꂽ�C�h�~�~�Y�n�[�W���^�DIchthy, Natural History of Fishes of Japan, 48: 33-40.
Ito, T., S. Matsui, K. Hirashima and R. Inui. 2025. Is Luciogobius dormitoris (Perciformes: Gobiidae) a valid species? Comparative morphology of two subterranean species of the genus Luciogobius. Journal of Asia-Pacific Biodiversity, corrected https://doi.org/10.1016/j.japb.2025.08.005 proof: (in press).
���ȁD2020�D���ȃ��b�h���X�g2020�̌��\�ɂ��āDhttps://www.env.go.jp/press/107905.html �i31 Mar. 2026�j�D
Kim, B. J. 2012. New record of a rare hypogean gobiid, Luciogobius pallidus from Jeju Island, Korea. Korean Journal of Ichthyology, 24: 306-310.
���}�����E�v�ؓc���l�E�{���_�V�D2020�D��Ŋ뜜���ރC�h�~�~�Y�n�[�̎������p����̏��߂Ă̋L�^�DNature of Kagoshima, 46: 267-269.
������㏼�D1955�D���ނ̌`�Ԃƌ��� II�D�菑�X�C�����D1605 pp.
������E�����I�P��E��{����E�I���I�E��q��D2026�D�X��������ނ܂� ��쒬���ӂ̐��������K�C�h�u�b�N�D�{���w�_�w������������_�w���猤���Z���^�[�����t�B�[���h�C�����D
���F�i�V��E�R��a�ƁE�O�R����Y�E���q����E���������Y�E���[�����D2025�D��錧�߉ϐ�͌���ō̏W���ꂽ����Ŋ뜜��C�h�~�~�Y�n�[��Q�i�n�[�ȁj�̎e���̋L�^�D��錧���R�����ٌ����C28: 73-79. LINK
OhmyNews. 2024. ���E�I�H�M��u�C�h�~�~�Y�n�[�v�C���ς��甭���D(In Korean) https://m.ohmynews.com/NWS_Web/Mobile/at_pg.aspx?CNTN_CD=A0003033003�i31 Mar. 2026�j�D
���������E�R�㗳���E�����O���E�b��ÍW�E�����L���D2024�D���m���ɂ�����C�h�~�~�Y�n�[��Q�̕��z�E��������ь`�ԓI�E��`�I�����DIchthy, Natural History of Fishes of Japan, 43: 20-37. LINK J-Stage
���������E�R�㗳���E���i ���E�k �F��E���c �āE�����O���E�b��ÍW�E�����L���D2026�D���m���ɂ�����C�h�~�~�Y�n�[�ƃh�E�N�c�~�~�Y�n�[�ߎ���̒lj��L�^�DAquatic Animals�C2026: AA2026-17.
���� ���D2002�D�C�h�~�~�Y�n�[�Cpp. 194-195�D���m�����b�h�f�[�^�u�b�N�m�����ҁn�ҏW�ψ���i�ҁj���m�����b�h�f�[�^�u�b�N�m�����ҁn�\���m���̐�ł̂�����̂���쐶�����\�D���m�������������ۑS�ہC���m�D
������P�E�䓡����E�� ����D2021�D�������암��3�͐�œ���ꂽ�C�h�~�~�Y�n�[�i�X�Y�L�ځF�n�[�ȁj�̋L�^�D�l�����R�j�Ȋw�����C14: 12-18. LINK
Regan, C. T. 1940. The fishes of the gobiid genus Luciogobius Gill. Annals and Magazine of Natural History (Series 11), 5: 462-465.
�a��_��E���V���G�E��؎��V�E���쒼�K�E�������l�D2019�D�É����Y�~�~�Y�n�[�����ނ̕��ފw�I�����i�\��j�D���C���R���C12: 29-96. LINK
�Ɛߖ�E�����@���D1976�D�C�h�~�~�Y�n�[ Luciogobius pallidus �̐��Ԃɂ��āD���a51�N�x���{���ފw�w��N��u���v�|�W�D���{���ފw��C�����D28 pp.
�����C�m��w���A��������D2025�D�}�^ �~�~�Y�n�[���Q�D�����C�m��w���A��������C�����D
Tomiyama, I. 1936. Gobiidae of Japan. Japanese Journal of Zoology, 7: 37-112.
�R�㗳���E���������E�����O���E�����L���D2023�D���m���ɂ�����n�����~�~�Y�n�[�����ނ̕��z�Ɛ����D���m��w���H�w���I�v��6���ʍ���1���C��114��y�������w����u���v�|�W�D
�R�����V��E�� �x�V��E�ɖ،��l�E�R��F���D2021�D�ΐ쌧����ѕ��Ɍ��̓��{�C���݂��瓾��ꂽ�C�h�~�~�Y�n�[�DIchthy, Natural History of Fishes of Japan, 10: 13-20.
�g�c���j�E���Ê�q�E�[�쌳���Y�E�{�ؗ��v�D2006�D���茧�呺�p�Y�C�h�~�~�Y�n�[O�^�CLuciogobius sp.�̐��ԁC�����j�Ǝ���D���茧�����w��C6: 113-125.
�ʐ^�W�{�F ��i�W���^�j�GBSKU 160132�C32.4 mm SL�C���m�����|�s �Ԗ��C2026�N3��28���C ���i�D���^�j�GBSKU 138743�C48.6 mm SL�C���m���{��s ���ː�C2025�N5��31���C �̏W�E�B�e�F���������E�k �F��D
�i���������j
2026�N2���̋�
![]()

�q���e���O�n�M Naso annulatus (Quoy and Gaimard, 1825)�i�j�U�_�C�ڃj�U�_�C�ȁj
�@ ����͐ߕ��ɂ��Ȃ�ŁC�����ɂȂ�Ɨ��h�ȋS�̊p�̂悤�Ɍ����闧�h�ȕ��������ɂ��ďЉ�܂��D���Ƃ͊���O���̕����̂��Ƃ������܂��i���V�E���R�C2013�j�D
�@�q���e���O�n�M Naso annulatus (Quoy and Gaimard, 1825)�͍g�C���܂ރC���h�E�����m�ɕ��z���C�����ł͏��}��������x�͘p�ȓ삩����L�^����Ă��܂��i��t�C2013�GGolani�C2025�j�DNaso �Ƃ��������̓��e����� natus�i�@�j�ɗR�����C���̓����I�ȊO�����\����Ă��܂��i���V�E�����C2015�j�D�포����annulatus �̓��e����Łu�`�́v�Ƃ����Ӗ��ł���C���h��ɔ��F�̗֏�͗l�����邱�Ƃ�w�h������h�C�������\�h�ɂ����Ĕ������ǂ��邱�ƂɗR������悤�ł��D�iQuoy and Gaimard, 1825�G���V�E�����C2015�j�D�`���ŕ����u�S�̊p�v�ƗႦ�܂������C�p��ł́uunicornfishes�v�ƌĂ�C�z��1�{�̊p�����`���̐������́u���j�R�[���v�ɂȂ��炦���Ă��܂��D�{��͔��h�̌㉏�������C��������{�E��������̑O���܂ł̒������قړ������C�w�h�� 5-6 �� 28-29 ����C�\�h��2�� 27-30 ����C���h�� 17-19 ����C���h��1��3����Ƃ��������������܂��i���c�C2013�j�D���̑��ɁC�Y�ł͔��h�̗��[������ɐL�т邱�Ƃ��ڂ������܂����C���h�̋e�̉Ԃт�̂悤�Ȗ͗l�������I�ł��i�g��E���\�C2023�j�D�����e���O�n�M���ł͐F�N�₩�Ȏ�����܂����C�q���e���O�n�M�͑S�̓I�ɊD�F�ł���C�ǂ���������������ۂ��܂��D�ʐ^�̌̂͑̒� 158.7 mm �ł��邽�߁C�����Ɍ�����p��ˋN�����B���Ă��܂���D��������̂͂��C�����͕����܂߂đS��1 m�ɂ��Ȃ邻���ł��iKuiter and Debelius, 2001�j�D�������ˋN�����q���e���O�n�M����݂��i�F�͂ǂ̂悤�ɉf���Ă���̂��C�����Ďז��ł͂Ȃ��̂��C�ɂȂ�Ƃ���ł��D�����������������������ۂɌ��Ă݂������̂ł��D�c���͒P�ƁC�܂��͏����ȌQ��Ő������܂����C���{���̑傫���ɐ������������́C��ʈ��T���S�ʈ�ŌQ��������ĕ�炵�Ă��܂��iKuiter and Debelius, 2001�G��t�C2013�j�D
�@�{��͎e�t���̕��V���������C�ꕔ�͓������m�̓��ׂɂ܂ł����U���܂��i���\�ق��C2013�j�D�j�U�_�C�Ȃ̗c���͓��������Ȃ��C���肪����ł��i���}�C2020�j�D�j�U�_�C��6����80�L����̂����C���̒t�����m���Ă���̂͑S�̂�10�������Ƃ����Ă��܂��iLeis�C2015; �Ë��E�{���D2024�j�D����ق��ɂ��ƁC�e���O�n�M���̒t�����j�U�_�C�ȑS�̂Ɠ��l�ɁC���{�Y�̖{��13��̂����C3��̗c����������݂̂ł��i����ق��C2014�j�D���ꂩ��̌����ŁC�{���̎e�t��������ɖ��炩�ɂȂ��Ă����̂��y���݂ł��D
�@�q���e���O�n�M�́C�e�t���̎���͂��Ȃ菬�����C�C�̗���ɐg��C���ĕY���Ă��܂����C��������ɂꎩ���̈ӎv�ŗ͋����j���܂��D��������̗���ɂƂ��ꂸ�C�����̈ӎv�������čs���ł���l�ɐ����������ł��D
�Q�l����
��t�@��D2013�D�j�U�_�C�ȁD�{���_�V�E�o�H�T��E�Óc�a�F�E���Y�\��i�ҁj�Cpp. 351-352�D���������O���� �������ƒ|���̋��ށD��������w�������������فC�������D
����@�āE�^��얤�E����M�a�D2014�D�j�U�_�C�ȁD���R�@�Y�i�ҁj�Cpp. 1348-1357�D���{�Y�t���}�ӑ��ŁC���C��w�o�ʼn�C�`��D
�Ë������E�{���_�V�D2024�D�����哇���瓾��ꂽ�W�{�Ɋ�Â����{���L�^�̃j�U�_�C�� ���� Acanthurus auranticavus �}�W�i�C�N���n�M�i�V�́j�D���ފw�G��, 71(2): 163�|171
Golan, D�D2025�DUpdate of Red Sea (Lessepsian) fish species in the Mediterranean Sea since the 2nd CIESM Atlas of Exotic Fish. Mediterranean Marine Science, 26(1):149-155.
���}�\���D2020�D�X�Y�L�ڃj�U�_�C��Acanthuridae�D���}�\���E���@���ˁE�R�c��F�E�{���_�V�i�ҁj�Cpp. 484-489�D��G�s�ꋛ�ސ}�ӁD��������w�������������فC�������D
Kuiter, R. H. and H. Debelius. 2001. Surgeonfishes, rabbitfishes and their relatives. TMC Publishing, Chorleywood. 208 pp.
Leis, J. M. 2015. Taxonomy and systematics of larval Indo-Pacific fishes: a review of progress since 1981. Ichthyological Research, 62: 9-28.
���V�O���E�����`�G�D2015�D���{�Y���ޑS��̊w�� �ꌹ�Ɖ���D���C��w�o�ŕ��C�`��D372 pp.
���V�O���E���R�k���D2013�D���ފT���D���V�O���i�ҁj�Cpp. 3-30�D���{�Y���ތ���.�S��̓���.��O�ŁD���C��w�o�ʼn�C�`��
Quoy, J. R. C. and J. P. Gaimard. 1825. Description des Poissons. Chapter IX. In: Freycinet, L. de, Voyage autour du Monde...exe�Lcute�L sur les corvettes de L. M. 192-401 [1-328 in 1824; 329-616 in 1825], Atlas pls. 43-65.
���\�@�G�E��h���F�E�ɓ����p�E�{���_�V�D2013�D���{���L�^�̃j�U�_�C�ȃe���O�n�M���̋H��}�T�J���e���O�n�M�i�V�́j�Ƃ��̕��z�����D�_�ސ쌧�������ٌ����i���R�Ȋw�j, 42 : 91-96.
���c�a�F�D2013�D�j�U�_�C��Acanthuridae�D���V�O���i�ҁj�Cpp. 1619-1631, 2215-2218�D���{�Y���ތ���.�S��̓���D��O�ŁD���C��w�o�ʼn�C�`��D
�g��Y��E���\�@�G�D2023�D�R�k�n���f�B�}��13 ������ ���{�̊C�����D�R�ƌk�J�ЁC�����D543pp.
���ڂ����R���j���N�̎s�ꋛ�L�ސ}�ӂ��q���e���O�n�M�̃y�[�W
�ʐ^�W�{�FBSKU 135777, 158.7 mm SL, 2024�N4��14��, ���m���y�������s�{�V�C��������`�C�ނ�C�̏W�ҁF�����y���D
�i�����y���j
2026�N1���̋�
 �@
�@
�A�Y�}�n�i�_�C Zalanthias azumanus (Jordan and Richardson, 1910)�i�y���J�ڃn�i�_�C�ȁj
�@���m��w���H�w�������Ȋw�ȁ@�C�m�����w�������z�[���y�[�W�������̊F�l�C�{�N���ǂ�����낵�����肢�������܂��D 1���͉��N�̗ǂ��g�������g�ɓZ�����������Љ�܂��D
�@��N�C�n�i�_�C�ȋ��ނ̒��ōő��̗L���퐔���܂�ł����C�Y�n�i�_�C�� Plectranthias Bleeker, 1873�́C��I�ȕ��q�n����͂���ɂ������ފw�I�Ȍ������s���C�]���̃C�Y�n�i�_�C����2�V�����܂�7���ɍו�����܂����iTang and Chen, 2025�j�D���̂����C�A�Y�}�n�i�_�C�� Zalanthias Jordan and Richardson, 1910 �͋��C�Y�n�i�_�C���̕W���ɉ����C���h���}�h�𐔂�8+7�C���h������㕔��1-2����������ĕ��}����C��_�o����3�{�C�̒��ɑ�����|�Ԓ����ʏ�5.0-6.3 %�iZalanthias kelloggi �ł�7.0-8.3 %�j�C�O�ҊW�������ɞ��������Ȃ��C���h�����������C�e�̊����ю������ɔ������т������ƂȂǂ̓����œ��ȑ����Ǝ��ʂ���܂��iTang and Chen, 2025�j�D
�@Tang and Chen (2025)�ɂ��ƁC�{���ɂ͓��{�ɕ��z����A�Y�}�n�i�_�C Zalanthias azumanus (Jordan and Richardson, 1910) ���͂��߁CZ. alleni (Randall, 1980), Z. bilaticlavia (Paulin and Roberts, 1987), Z. exsul (Heemstra and Anderson, 1983), Z. kelloggi (Jordan and Evermann, 1903), Z. maculicauda (Regan, 1914), Z. melanensis (Randall, 1980), Z. nazcae (Anderson, 2008), Z. parini (Anderson and Randall, 1991) ��9�L���킪�܂܂�܂��D���̂����C�A�Y�}�n�i�_�C�͌v���`����������i�Ȃǂ̓����� Z. kelloggi�CZ. maculicauda�C����� Z. melanensis �ɗގ����܂����C�����L�E�ؐ���33-37�ivs. Z. maculicauda �ł�32-35�CZ. melanensis �ł� 32-34�j�C�����̉��{�������iZ. maculicauda �ł͖��j�C�̑��ʒ��ԕ��̈Əт̉��[���\�h���̉���� 2-3 ��������ɒB����iZ. kelloggi �� Z. maculicauda�ł͑̑��ʂ̒����t�߂܂ŁCZ. melanesius �ł͕������ɒB����j�C�������ɏc��� 3-5�����̕��ŁC������������艡��� 1-2���������܂ŒB����F�f�E�����W�����Ə�������iZ. kelloggi �ł͕����c��� 5-6�����ŁA�����������Ɉ�v����Ə���CZ. maculicauda �ł͔����������ɏc���ŁA����� 3-5�����̕������ȉ~�`���CZ. melanesius �ł͔����̔w�����畠���ɒB���鉡�сj�����Ȃǂŗe�ՂɎ��ʂ���܂��iGill et al., 2021; �a�c�ق��C2022�j�D
�@�A�Y�}�n�i�_�C�́CSteindachner and Doderlein (1883) �́u���{�̋��ނɊւ���m���ւ̍v���v�i�h�C�c��j�̒��ŁC�����p�Y�̐���������̕W�{�Ɋ�Â��C���[�g���B�b�q�E�f�[�f�����C���iLudwig H. P. Doderlein�C1855-1936�j�ɂ��Anthias japonicus Doderlein in Steindachner and Doderlein, 1883 �Ƃ��ċL�ڂ���܂����DDoderlein�͖�������ɂ��ق��O���l���t�Ƃ��ė��������h�C�c�l�̐����w�҂ŁC1879�N����1881�N�܂œ����鍑��w��w���Ɍق��ē��{�ɑ؍݂��Ă��܂����D�������C���̌� Jordan and Richardson (1910)�́CAnthias japonicus Doderlein in Steindachner and Doderlein, 1883 �� A. japonicus Bloch, 1793 [���݂̃��R�V�}�^�}�K�V���� Scolopsis �� Scolopsis vosmeri (Bloch, 1792)] �ɂ���Đ�悳��Ă������Ƃ��w�E���CPseudanthias azumanus �̒u��������܂����D����ɁCJordan and Richardson (1910)�� Anthias kelloggi Jordan and Evermann, 1903 ���^�C�v��Ƃ��āC�����������m���ݗ����� Zalanthias �������̃����N�Ƃ��āC�{�������Ɋ܂߂����߁C�w���� Pseudanthias (Zalanthias) azumanus Jordan and Richardson, 1910 �ƂȂ�܂����D���̌�CKatayama (1959, 1960) �� Zalanthias ��Ɨ����Ƃ��Ĉ����CZalanthias azumanus �Ƃ��܂����D����ɂ��̌�CRandall (1980) �͋��C�Y�n�i�_�C�� Plectranthias Bleeker, 1873 �̕�I�ȕ��ފw�I�Č������s���CZalanthias �� Plectranthias �̐V�Q�ٖ��Ƃ��������łȂ��CP. azumanus ���C�n���C�������^�C�v�Y�n�Ƃ��� P. kelloggi �Ɠ����Ƃ݂Ȃ��C�V�Q�ٖ��Ƃ��܂����D�܂��C���҂̍��ق͈���Ƃ��Ĉ�������͈͂ɂ���ƍl���CPlectranthias kelloggi azumanus (Jordan and Richardson, 1910) �Ƃ��܂����D���̌�C���\�i2013�j�Ȃǂł́C����Ƃ��Ď�舵���Ă��܂��D����CRandall (1996) �� P. azumanus �� P. kelloggi �̐V�Q�ٖ��Ƃ݂Ȃ��CKuiter (2004) �� P. azumanus ��L����Ƃ��Ĉ����ȂǁC�w���̈����͕K���������ꌩ���������Ă��܂���ł����D�������CGill et al. (2021)�� P. kelloggi �� P. azumanus ���O���`�ԂƑ̐F�ɂ�薾�ĂɎ��ʂł���Ƃ��āC���҂�L����Ƃ��Ĉ����܂����D������C�����E�{���i2022�j��a�c�ق��i2022�j�Ȃǂ̋ߔN�̕ł��C����͂��ꂼ��L����Ƃ���Ă��܂��D�����āCTang and Chen (2025)�ɂ��Zalanthias �͍ĂїL�����Ƃ��ĕ����������߁C�A�Y�}�n�i�_�C�ɓK�p�����w���� Zalanthias azumanus �ƂȂ�܂����D
�@�A�Y�}�n�i�_�C�͓��{�C�؍��C��p�C�I�[�X�g�����A�C����уx�g�i���������Ă��܂��i�a�c�ق��C2022; Tang et al., 2025�j�D����C���{�ߊC�ɂ����ẮC��錧����{�茧�ɂ����Ă̖{�B�|��B�����m���݁C�H�c�����璷�茧�ɂ����Ă̖{�B-��B���{�C���݂��͂��߁C�F���������݁C�������p�C�V�������n���C�B���ߊC�C���茧�Δn�C�ܓ��C���������F������s�����̑铇�C����������i�Ǖ����C���v�V�]�����܂ރg�J���C�����Q�������哇�C���ꏔ���C���d�R�����C�������C��E�����C�R�C��B�|�p���I�C��C����M��C�~�C����ѓ��V�i�嗤�I���ӕ�����L�^������܂��i�a�c�ق��C2022�j�D�{��͒ʏ퐅�[ 60-280 m �ɐ�������Ƃ���iGill et al., 2021�j�C�a�c�ق��i2022�j�͓��{�ߊC�ɂ������I�ȕ��z�Ɛ����Ƃ̊W����C�{�킪���� 8-18 ���͈̔͂ɓK���\�Ɛ������Ă��܂��D���{�����̑����m����ѓ��V�i�C�ł́C��r�I���ܓx�Ɉʒu���鑊�͘p��x�͘p�ɂ����āC1-3 ���̒ᐅ�����ɐ��[ 30-31 m �Ŋώ@����Ă���ق��C���m�����̓��ߊC�ŋL�^���ꂽ���[ 400 m �������ƁC��r�I��ܓx�̉��ꌧ�ɍ]�����ł͐��[ 330-350 m�C����ɒ�ܓx�Ɉʒu�����p�ł͐��[ 400 m ����̏W����Ă��܂��D���̂悤�ɁC�{��͒�ܓx�C��قǐ������[���[���Ȃ�Ƃ��������ȌX�����m���Ă��܂��D
�@�A�Y�}�n�i�_�C�͕W���̒����� 120 mm ���x�̏��^���ł͂���܂����C�×~�ɉa��ߐH���鐫���������߁C�g���C���C�g�]�[���ōs����[�C�����ނ�ɂ����āC�������Q�X�g�Ƃ��ďo����Ƃ���������܂��D�܂��C�����ߋ��ɖK�₵����p�k���̎s��ł́C�{��Ɠ��l�ɋ��C�Y�n�i�_�C���Ɋ܂܂�Ă����C�Y�n�i�_�C Plectranthias kamii Randall, 1980�C�L�I�r�C�Y�n�i�_�C Plectranthias sheni Chen and Shao, 2002�C������ Plectranthias purpuralepis Tang et al., 2020 �ȂǂƂ܂Ƃ߂Ĕ̔�����Ă���l�q��ڂɂ��C���ۂɍw�����܂����i�ʐ^�j�D�܂��C���̂悤�ɔ̔�����Ă��闝�R�Ƃ��āC���n�ł́C�^�C�Ȃ�L���g�L�_�C�ȁC�A�}�_�C�ȋ��ނ�ΏۂƂ��������ނ�ɂ���č��l����Ă���ƌ��n�̕�����f���܂����D
�@�{��͉��a�Ȑ��i�ɉ����C����g�n�`���D�ރn�i�_�C�ȋ��ނł��邱�Ƃ���C��ʈ�ɐ�������Ƃ�����ۂ������������Ǝv���܂��D�������C�{�킪���I�ꂩ����̏W����邱�Ƃ́C���܂�m���Ă��Ȃ���������܂���i�R�c�C1986�j�D���ۂɁC�y���p�̍��I��ő��Ƃ������J�苙�i������g�ԋ��j�ł����l����܂��D���m���Ő��g�����ꂽ�A�Y�}�n�i�_�C�́C�q�������ނ�z�E�{�E�ȋ��ނƂƂ��Ɂu�V�Ղ�i���̂���g��p�������ܗg���j�v�։��H����C�ӎ����Ă��Ȃ��Ƃ��C���m���œV�Ղ�����ɂ������Ƃ̂�����́C�ԐړI�ɃA�Y�}�n�i�_�C�𖡂���Ă���̂�������܂���ˁD
���p����
Gill, A. C., J. J. Pogonoski, G. I. Moore and J. W. Johnson. 2021. Review of Australian species of Plectranthias Bleeker and Selenanthias Tanaka (Teleostei: Serranidae: Anthiadinae), with descriptions of four new species. Zootaxa, 4918: 1-116.
Jordan, D. S. and R. E. Richardson. 1910. A review of the Serranidae or sea bass of Japan. Proc. U.S. Natl. Mus., 37: 421-474.
Katayama, M. 1959. Studies on the serranid fishes of Japan (I). Bull. Fac. Educ. Yamaguchi Univ., 8: 103-180.
Katayama, M. 1960. Fauna Japonica. Serranidae (Pisces). Biogeographical Society of Japan, Tokyo. i-viii + 1-189 pp., pls. 1-86.
Kuiter, R. H. 2004. Basslets, hamlets and their relatives: a comprehensive guide to selected Serranidae and Plesiopidae.TMC Publishing, Chorleywood. 216 pp.
Randall, J. E. 1980. Revision of the fish genus Plectranthias (Serranidae: Anthiinae) with descriptions of 13 new species. Micronesica, 16: 101-187.
Randall, J. E. 1996. Two new anthiine fishes of the genus Plectranthias (Perciformes: Serranidae), with a key to the species. Micronesica, 29: 113-131.
���\�@�G�D2013�D�n�^�ȁD���V�O��(��)�Cpp. 757^ 802, 1960^1971.���{�Y���ތ��� �S��̓��� ��O��.���C��w�o�ʼn�C�`��.
Steindachner, F. and L. Doderlein. 1883a. Beitra�Nge zur Kenntniss der Fische Japans (I.). Anz. Kaiserl. Akad. Wiss. Wien, Math.-Naturwiss. Kl., 20(7): 49-50.
Tang, C.-N. and W.-J. Chen. 2025. A 40-year taxonomic enigma: multigene phylogeny resolves the polyphyly of Plectranthias (Perciformes: Anthiadidae) and supports a revised taxonomy. Zool. J. Linn. Soc., 205(3): zlaf148.
Tang, C.-N., Q. V. Vo and A. C. Gill. 2025. Sayonara flavolineata (Perciformes: Anthiadidae), a new deep-water anthiadid perchlet from Vietnam. Raffles Bull. Zool., 73: 698-708.
�R�c�~�F�D1986�D�A�Y�}�n�i�_�C�Cp. 144�D�R�c�~�F�E�c�� ���E�ݓc���O�E�{��N���i�ҁj���V�i�C�E���C�̋��ށD���Y�����C�搅�Y�������C����D
�a�c�p�q�E�O�엲���E�b��ÍW�E�{���_�V�D2022�D�R�`���C�������p����щ����哇����̃A�Y�}�n�i�_�C�i�n�i�_�C�ȁj�̐V���ȕ��z�L�^�C����і{��̒n���I���z�Ɛ������[�E�����̊֘A���DIchthy, Natural History of Fishes of Japan, 24: 24-32.
�ʐ^�W�{�FBSKU 137637, 98.3 mm SL, 2024�N11��15���C���m�����m�s��������`�C���J��ԁC�i�ہC���|�s���C���[130-140 m�D�̏W�ҁF����D���Y�C�V���\��C��㊰���Y�C����D
�i����j
|
Copyright (C) Laboratory
of Marine Biology, Faculty of Science, Kochi University (BSKU) |
{kind=link}